Химия, Биология, подготовка к ГИА и ЕГЭ. Самые древние бактерии архебактерии
Архебактерии | Дистанционные уроки 27-Май-2014 | Нет комментариев | Лолита Окольнова или Археи — группа древних прокариотических микроорганизмов, выделяемых в отдельный домен 
Ранее археи объединяли с бактериями в общую группу, называемую прокариоты , и они назывались архебактерии, однако сейчас такая классификация считается устаревшей: установлено, что археи имеют свою независимую эволюционную историю и характеризуются многими биохимическими особенностями, отличающими их от других форм жизни. Название «Археи» произошло от периода Земли, в котором эти организмы были доминирующими — Архейской эры, но в школьном курсе чаще встречается название «Архебактерии». Существуют 3 группы архебактерий: 1- метанобразующие -окисляют углекислый газ, 2 – галобактерии — это «экстремалы» — они выдерживают такие условия, как, например, концентрированные растворы соли (Мертвое море) Они могут жить как за счет органических субстратов (гетеротрофные аэробы), так и за счет энергии Солнца, имея в своих мембранах светочувствительный пигмент –бактериородопсин (фотоавтотрофы). 3 – экстремальные термофилы — это, соответственно, температурные «экстремалы» — их среда жизни — термальные источники. Температура около 100 °С для них не проблема Они могут использовать для питания как органические соединения (аэробные гетеротрофы), так и неорганические соединения (аэробные и анаэробные хемотрофы) У архебактерий есть черты, роднящие их с бактериями, но есть так же общие признаки с эукариотами: | Сходства архебактерий и бактерий | Сходства архебактерий и эукариот | Свойства только арбактерий | | нет ядра нет мембранных органелл | нет муреина | архебактерии содержат псевдомуреин | | кольцевая ДНК — нуклеойд | процессы транскрипции и трансляции схожи | в составе мембраны есть липиды генетически отличаются от эукариот и бактерий | | размер и форма клеток есть жгутик | | форма клетки может быть квадратной однослойная мембрана | | некоторые похожие обменные процессы | Хемоавтотрофы Архебактерии используют значительно больше источников энергии, чем эукариоты: начиная от обыкновенных органических соединений, таких как сахара, и заканчивая аммиаком, ионами металлов и даже водородом. | | размножение: 1) бесполое надвое 2) почкованием 3) «половое» — коньюгация | | не образуют спор |
- именно архебактерии образовали запасы природного газа (в частности метана) на Земле
- удивительно и то, что среди архебактерийнет паразитов и болезнетворных форм, что заметно отличает их от бактерий.
 Большой вклад в изучение строения, эволюции архебактерий внес академик РАН Б.В.Громов, профессор Санкт-Петербургского университета. Им был создан уникальный банк архебактерий. Филогенез, экология, систематика архебактерий были досконально изучены чл. – корр РАН Г.А. Заварзиным. В своих многочисленных экспедициях на Камчатку, другие экстремальные районы Земли, им были обнаружены и описаны многие виды и группы архей и эубактерий. Еще на эту тему: Обсуждение: "Архебактерии" (Правила комментирования) distant-lessons.ru Археи | Info-Farm.RU Археи (лат. Archaea, от греч. Αρχαία — «старые», также архебактерии — Archaebacteria) — одна из групп живых организмов, в которую входят микроскопические одноклеточные прокариоты, очень отличаются рядом физиолого-биохимических признаков от настоящих бактерий (эубактерий). Хотя до сих пор является неопределенность в точном филогенезе этих групп, археи, эукариоты и бактерии являются фундаментальными группами в так называемой системе трех доменов. Подобно бактерий, археи — это одноклеточные организмы, не имеющие ядра и поэтому классифицируются как прокариоты (Prokaryota) — известные также как Monera в таксономии пяти царств. Археи сначала было обнаружено в экстремальных средах, но затем они были найдены во всех типах экосистем. Архебактерии существенно отличаются от других микроорганизмов по составу и последовательностью нуклеотидов в рибосомных и транспортных РНК. История Археи было идентифицировано 1977 Карл Вёзе (Carl Woese) и Джорджем Фоксом (George Fox), основываясь на их отличия от других прокариот (филогенетические деревья, основанные на 16S рРНК). Когда 1977 биолог Карл Вёзе изучал генетику микробов, производили метан, он понял, что они отличаются от всех известных бактерий. Стенка клетки была уникальной — производила уникальные энзимы и ее генетический ряд не был похож ни на что исследуемое раньше. Очень быстро — в течение часа — стало понятно, что есть еще что-то, третья группа. Это был момент открытия. Карл Вёзе открыл третью форму жизни — группу одноклеточных, которую он назвал археями. Он открыл форму жизни, которая способна существовать где угодно, включая самые невероятные места. Эта группа была сначала назван археобактериямы (Archaebacteria) в отличие от эубактерий (Eubacteria), и они рассматривались как царства или подцарства. Вёзе убеждал, что археи являются представителями совсем другой ветви живых существ. Он позже переименовал группы в архей и бактерий, чтобы подчеркнуть это, и аргументировал тем, что вместе с эукариотами они охватывают три группы живых организмов. Биологический термин архея (археи) не следует путать с архейская геологической эрой, также известной как Археозойська эра. Последний срок ссылается на древний период земной истории, когда археи и бактерии были единственными клеточными организмами, живших на планете. Возможные окаменелости этих микробов были датированы почти 3800000000 лет назад. Археи, бактерии и эукариоты Археи подобные других прокариот в большинстве аспектов структуры клетки и метаболизма. Однако, их генетическая транскрипция и трансляция — два центральных процессы в молекулярной биологии — не проявляют типичных для бактерий особенностей, но чрезвычайно подобные этих процессов у эукариот. Например, система трансляции архей использует эукариотические факторы инициации и элонгации, а их система транскрипции применяет TATA-связываемые белки и TFIIB, как у эукариот. Гены тРНК и рРНК археи содержат уникальные археальни интроны, которые не похожи ни к эукариотических, ни к бактериальным интронов. Некоторые другие характеристики также выделяют архей. Подобно бактерий и эукариот, археи имеют основанные на глицерине фосфолипиды. Однако, три особенности липидов архей необычны: - Липиды архей уникальны, так как стереохимия глицерина — обратная той, что найдена в бактериях и эукариот. Это убедительное свидетельство другого биосинтетического пути.
- Большинство бактерий и эукариот имеют мембраны, составленные преимущественно из эфир-липидов глицерина, тогда как археи имеют мембраны, состоящие из эфир-липидов глицерина. Даже когда бактерии имеют эфир-связанные липиды, стереохимия глицерина — бактериальная форма. Эти различия, возможно, является адаптацией части архей к гипертермофилии. Однако, стоит отметить, что даже мезофилични археи имеют эфир-связанные липиды.
- Липиды архей основанные на изопреноидного боковом цепочке. Это пятиуглеродный химическая группа, также обычная для каучука и является компонентом некоторых витаминов, распространенная в бактериях и эукариот. Однако, только археи применяют эти вещества в своих клеточных липидах, часто как боковые цепочки C-20 (четыре мономера) или C-40 (восемь мономеров). В некоторых архей изопреноидного боковой цепочку C-40 — фактически достаточно длинный, чтобы пройти сквозь мембрану, формируя монослой вместо клеточной мембраны с группами фосфата глицерина на обоих концах.
Эта адаптация распространенная в чрезвычайно теплолюбивых археи. Хотя они и не уникальны, клеточные стенки архей также необычны. Например, клеточные стенки большинства архей образованные внешними слоями белков или S-слоем. S-слоя распространенные в бактериях, они служат единственным компонентом клеточной стенки в некоторых организмах (например, в Planctomyces) или внешний слой во многих организмах с пептидогликана. За исключением одной группы метаногенов, археи не имеют пептидогликановый стенки. Однако даже в этом случае, пептидогликаны очень отличаются от разновидности, найденного в бактериях. Археи также имеют жгутики, которые значительно отличаются по составу от очень похожих жгутиков бактерий. Бактериальные жгутики — это измененная система секреции III типа, тогда как жгутики архей напоминают ворсинки IV типа (type IV pili), которые используют sec-зависимую систему секреции, несколько подобную, но все же отличную от системы секреции II типа. Среда Многие архей — экстремофилы. Некоторые живут при очень высоких температурах, часто выше 100 ° C, как те, которых нашли в гейзерах и черных курильщиках. Других найдено в очень холодных средах или в чрезвычайно соленой, кислой или щелочной воде. Однако, некоторые археи — мезофилы, живут в средах, подобных болота, сточных вод и почвы. Многие метаногенных архей найдено в пищеварительных трактах животных, например, жвачных животных, термитов и людей. Археи не патогенны, и неизвестно, чтобы какие-либо из них вызвали болезнь. Архей конечно разделяют на три группы, в соответствии с среды. Это — галофиты, метаногены и термофилы. Галофилы живут в чрезвычайно соленых окружающих средах (например, многие из них живут в Мертвом море и на юге залива Сан-Франциско, придавая ей ярких цветов: от красного до зеленого). Метаногены живут в анаэробных окружающих средах и производят метан. Их можно найти в отложениях или в кишечниках животных. Термофилы живут в местах с высокими температурами, как, например, горячие источники. Эти группы не обязательно согласуются с молекулярным филогенезом, не полные но не взаимоисключающие. Тем не менее, они — полезный начальный пункт для подробного изучения. Недавно несколько исследований показали, что археи существуют не только в мезофильных окружающих средах, но также иногда присутствуют в значительном количестве при низких температурах. Становится все более принятым, что метаногены обычно присутствуют в низкотемпературных средах, как, например, холодные осадки. Некоторые исследования даже свидетельствовали, что в этих температур путь метаногенеза может изменяться благодаря термодинамическим ограничениям, обусловленным низкими температурами. Возможно, даже, важнее то, что большое количество архей найдено в большинстве океанов, в холодных средах (Giovannoni и Stingl, 2005). Эти археи, которые относятся к группам, ранее исследованных, могут случаться в чрезвычайно высоких количествах (до 40% микробной биомассы), хотя они не были изолированы в чистой культуре. Сейчас мы не имеем почти никакой информации о физиологии этих организмов, что означает, что их влияние на глобальные биогеохимические циклы неизвестен. Одно недавнее исследование (Könneke и другие, 2006) показало, однако, что одна группа морских Crenarchaeota способна к фиксации азота, а это — черта, до сих пор неизвестна среди архей. Генетика Как правило, археи имеют одиночную кольцевую хромосому, размер которой может достигать 5751492 пар нуклеотидов в Methanosarcina acetivorans, что самый известный геном среди архей. Одну десятую размера этого генома составляет геном с 490885 парами нуклеотидов в Nanoarchaeum equitans, что маленький геном среди архей; он содержит только 537 генов, кодирующих белки. Также в архей обнаружены более мелкие независимые молекулы ДНК, так называемые плазмиды. Вероятно, плазмиды могут передаваться между клетками при физическом контакте, во время процесса похожего на конъюгации бактерий. Археи могут быть поражены вирусами, содержащими двухцепочечной ДНК. Эти виды не родственные с другими группами вирусов и имеют различные необычные формы, так как бутылки, крючки, капли и т. Эти вирусы были тщательно изучены на термофилы, в основном рядов Sulfolobales и Thermoproteales. В 2009 году был открыт вирус, содержащий одноцепочечную ДНК и поражает галофильные археи. Защитные реакции архей против вирусов могут включать механизмы близок к РНК-интерференции эукариот. Археи генетически отличны от эукариот и бактерий, причем до 15% белков, кодируемых одним геном бактерии, являются уникальными для этого домена, хотя функции многих этих белков неизвестны. Большая часть уникальных белков, функция которых известна, принадлежит евриархеотам и задействована в метаногенеза. Белки, общие для архей, бактерий и эукариот, участвующих в основных клеточных функциях и касаются в основном транскрипции, трансляции и метаболизма нуклеотидов. К другим различий архей можно отнести организацию генов, выполняющих связанные функции (например, гены, отвечающие за различные этапы одного и того же метаболического процесса), в оперон и большие различия в строении тРНК и их аминоацил-тРНК синтетаз. Транскрипция и трансляция архей больше напоминают эти процессы в клетках эукариот, чем бактерий, причем РНК-полимераза и рибосомы архей очень близки к аналогичным структур у эукариот. Хотя у архей есть только один тип РНК-полимеразы, по строению и функции она близка к РНК-полимеразы II эукариот, при этом подобные группы белков (главные факторы транскрипции) обеспечивают связывание РНК-полимеразы с промотором гена. в то же время другие факторы транскрипции архей более близки к таковым у бактерий. Процессинг РНК в архей проще, чем у эукариот, поскольку большинство генов не содержит интронов, хотя в генах их тРНК и рРНК их достаточно много, также они присутствуют в небольшом количестве генов, кодирующих белки. Размножение Археи размножаются бесполым путем: бинарным или множественным делением, фрагментацией или почкованием. Мейоза не происходит, поэтому даже у представителей конкретного вида архей существуют в более чем одной форме, все они имеют одинаковый генетический материал. Клеточное деление определяется клеточным циклом: после того, как хромосома реплицировать и две дочерние хромосомы разошлись, клетка делится. Детали были изучены только для рода Sulfolobus, но особенности его цикла очень схожи с особенностями у эукариот и у бактерий. Репликация хромосом начинается с многих точек начала репликации с помощью ДНК-полимеразы, похожей на аналогичные ферменты эукариот. Однако белки, управляющие клеточным делением, такие как FtsZ, которые формируют сжимающее кольцо вокруг клетки и компоненты септы, проходящей через центр клетки, схожие с их бактериальными эквивалентами. Археи не образуют споры. Некоторые виды Haloarchaea могут претерпевать изменения фенотипа и существовать как клетки нескольких различных типов, включая толстостенными клетками, устойчивыми к осмотического шока, позволяющие археям выживать в воде с низкой концентрацией соли. Однако эти структуры не служат для размножения, а скорее помогают археям осваивать новые среды обитания. Эволюция и классификация Название говорит о том, что эти организмы в наше время многими исследователями считаются древнейшими живыми организмами на Земле. Археи разделены на две основные группы, основанные на деревьях 16S рРНК: Euryarchaeota и Crenarchaeota. Две другие группы были также созданы для определенных экологических образцов и особого вида Nanoarchaeum equitans, найденных в 2002 Карлом Стеттером (Karl Stetter), но их сходство с других групп все еще исследовано очень плохо. Карл Вёзе (Carl Woese) убедил, что бактерии, археи и эукариоты представляют собой уникальные наследственные линии, которые довольно рано отклонились от наследственного Прогенота (Progenote) с плохо развитым генетическим аппаратом. Эта гипотеза отражена в имени археи (Archaea, от Греческого archae или древний). Позже он формально рассматривал эти группы как империи, каждая из которых охватывает несколько царств. Этот раздел стал самым популярным, хотя идея Прогенота не всеми поддерживается. Некоторые биологи, однако, утверждают, что, археобактерии и эукариоты произошли от специализированных бактерий. Взаимоотношения между археями и эукариотами остаются важной проблемой. Учитывая сходство, указанную выше, много генетических деревьев располагают эти две группы вместе. Некоторые ставят эукариот ближе к Eurarchaeota, чем к Crenarchaeota, хотя мембранная химия предлагает наоборот. Однако открытие архея-подобных генов в определенных бактериях, например, Thermotoga, делает их взаимоотношения трудным для определения. Некоторые исследователи считают, что эукариоты возникли из смешения археи и бактерии, они стали ядром и цитоплазмой, объясняют различную генетическую схожесть, но эта теория не может объяснить происхождение клеточных структур. Внутренняя классификация Архебактерии разнообразные по типу обмена веществ, физиологическими и экологическими особенностями: среди них встречаются аэробных и анаэробных, хемогетеротрофы и хемоавтотрофы, нейтрофилы и ацидофилы. Некоторым Архебактерии (галобактерии) присущ особый тип фотосинтеза, когда свет поглощается НЕ хлорофиллом, а бактериородопсинном. Другим археям властвуй энергетический процесс, в результате которого образуется метан. Описано более 40 видов археи (25 родов), относящихся к 5 различных групп: Метанобразующие бактерии, сиркоокислюючи термоацидофилы, сирковидновлюючи термофилы, галобактерии, термоплазма. Расшифровка одиночных генов привела к расшифровке целых геномов, сейчас 24 архейни геномы завершения и 22 частично завершено (1). Изображения по теме info-farm.ru Археи: кто они такие? Как известно, происхождение жизни и дальнейшая эволюция
живого на нашей планете объясняется учёными в основном пребиотической
(химической) эволюцией, в ходе которой органические вещества возникли из
неорганических молекул. Согласно последним исследованиям жизнь на Земле
появилась примерно 3,9 миллиардов лет назад, а примерно 3,5-3,8 миллиардов лет
назад существовал последний общий предок, от которого произошли бактерии и археи, а также эукариоты. Археи – это одноклеточные простейшие микроорганизмы, не
различимые невооружённым человеческим взглядом, которые не имеют ни ядра, ни
мембранных органелл (т.е. каких-либо постоянных структур, каждая из которых
выполняет определённые функции). Интересно, что до 1977 года учёные вообще не
разделяли царства археев и бактерий меж собой, как это сделано сейчас: археи
тогда назывались архебактериями, т.к. считалось, что в целом эти организмы
сходны между собой по многим признакам. В настоящее время учёные разделяют 4
царства – царство археев, царство вирусов, царство бактерий и царство
эукариотов (сюда относятся царства грибов, растений, животных и пр.).
Разделение всех организмов по царствам происходило в течение XIX-XX веков с развитием микроскопов и
продолжается по сей день. Бактерии и археи были отнесены в одно царство прокариотов по
многим признакам, первый, который обнаружили учёные – это поразительное сходство
по размеру и форме клеток. Несмотря на это, археи склонны принимать более
причудливые формы (например, квадратные), в то время как бактерии – правильные.
Учёные отмечают, что археи имеют и общее с эукариотами, например, при процессах
транскрипции и трансляции принимают участие одни и те же ферменты. По сравнению
с другими организмами отличительными чертами являются и транспортные РНК
(рибонуклеиновые кислоты, задача которых – транспортировка аминокислот к местам
синтеза белка). Размножаются археи бесполым путём: либо почкованием, либо
бинарным делением, в ходе которого расходятся в разные стороны клетки две
дочерние хромосомы, а после происходит непосредственно деление клетки на две
новых. При этом археи не способны образовывать споры. Учёные отмечают, что
редкие виды археев могут претерпевать изменение фенотипа (совокупности
характеристик особи) на протяжении жизненного цикла. Помимо этого, археи могут
существовать как клетки нескольких типов. Скорее всего, такая склонность
обуславливает способность приспосабливаться к самым экстремальным условиям
жизни. Явное
отличие же от эукариотов заключается в том, что археи способны использовать
гораздо больше источников энергии, их приспособленность к окружающей среде в
разы больше. Общее процентное их содержание от всей биомассы составляет
примерно 20%.Существуют
солеустойчивые археи, которые используют солнечные свет в качестве энергии, а
обитают в морях с высоким содержанием соли, например, в Мёртвом море. Учёными были найдены археи, которые в качестве источника
энергии используют аммиак и другие группы углеводородов, водород и ионы
металлов. Кстати, ещё одно отличие археев от эукариотов – они не многофункциональны,
как, например, сине-зелёные водоросли, которые способны получать энергию двумя
способами одновременно. Высокая приспособленческая способность археев изначально
заставила учёных отнести их к группе экстремофилов – т.е. к особой группе организмов, способных
размножаться и жить в каких-то экстремальных условиях (к таковым относятся горячие
вулканические источники, вечные льды, полярные моря и т.п.). Однако в
середине-конце XX века
стало всё больше находок их и в океане (в основном, на океаническом дне), и в болотах, и в засушливых районах, и в полярных морях, и
даже в самом человеке (в основном, они находятся в желудке, чаще всего - в толстой
кишке, для них это практически что идеальная среда размножения и жизнедеятельности). Неподдельный интерес к этим организмам вызывает тот факт, что археи способны осуществлять
некоторые сложные биохимические процессы, которые не под силу другим организмам:
в частности они могут вырабатывать метан или серную кислоту в процессе своей
жизнедеятельности. Без деятельности организмов царства археев невозможное
нормальное функционирование всей экосистемы (т.е. единства живого) и
круговорота углерода в природе. Археи способны удалять азот из экосистемы,
перерабатывая его в ходе своей деятельности, а также окислять аммиак, в результате
чего происходит образование нитритов, которые затем будут переработаны другими
организмами в нитраты, которые в свою очередь будут потреблены растениями.
Однако такая деятельность археев может сыграть и вредную роль для всего живого
на планете: упомянутое выделение метана в процессе жизнедеятельности играет на
руку возникновению парникового эффекта. Это объясняется с химической точки
зрения тем, что метан имеет большую склонность (в 25 раз выше, чем у
углекислого газа) к созданию этого эффекта. Археи могут принимать участие в явлении мутуализма – явлении
взаимовыгодного сожительства двух разных организмов, при котором выгоду от
сожительства получает каждая особь. Отличным примером является взаимодействие
простейших и археев в желудке у термитов. Простейшие, находящиеся вместе с
археями в желудке, способны разлагать поступающую в пищеварительный тракт
термита целлюлозу. При этом процессе выделяется достаточно большое количество
водорода, что способствует меньшему поглощению энергии простейшими. Здесь на
помощь приходят археи, которые потребляют выделяющийся водород в качестве
энергии для себя, выделяя при этом метан, выводящийся термитом наружу. Всё большие исследования царства археев доказывают, что их
роль в технологии и промышленном производстве поистине велика. Как уже
упоминалось выше, археи-экстремофилы являются отличными источниками некоторых ферментов,
которые способны продолжать функционировать при температурах, превышающих 100
°C. Эти ферменты используются при производстве некоторых продуктов питания с
пониженным содержанием лактозы – молока или сыворотки. Последние исследования учёных показывают, что в будущем
археи способны дать медицине новые сильнейшие антибактериальные средства,
потенциально лучшие по своему воздействию на организм современных. Единственное,
что не даёт права учёным испытывать их на подопытных животных – такие
антибиотики отличаются по своей структуре от привычных бактериальных, а
возможные реакции организма эукариотов с ними пока до конца не просчитаны. www.8lap.ru АРХЕБАКТЕРИИ И ЭУБАКТЕРИИ — Мегаобучалка В 1977 году К.Вёзе и Фоксом на основании сравнения последовательностей нуклеотидов в рРНК малой субъединицы рибосомы (16S-рРНК у прокариот и 18S-рРНК у эукариот) было выделено три, а не две группы организмов, которым присвоен ранг домена, этот таксон более высокий по отношению к царству: Bacteria, Archaea, Eukarya. К Bacteria относят истинные бактерии – эубактерии (например, E. coli), ранее обозначали как Eubacteria.К Archaea относят архебактерии (археи) - это крупная группа прокариот, распространенных преимущественно в экстремальных условиях, эволюционно наиболее древние организмы. Ранее обозначали как Archaebacteria. Третья группа организмов - эукариоты, ранее обозначали как Eukaryota. Поскольку концепция домена до сих пор дискутируется, термины эубактерии и архебактерии используются наряду с терминами археи и бактерии. Считается, что домены Archaea и Eukarya имеют общее происхождение и являются сестринскими таксонами, тогда как домен Bacteria представляет собой отдельную эволюционную ветвь, которая ответвилась раньше от общего корня. Общий предок трех основных типов жизни назван прогенотом. Представители домена Bacteria явились предками хлоропластов и митохондрий. Эукариоты произошли от прокариотического предка, который, возможно, был более родственен Archaea, чем Bacteria. Прокариоты не представляют собой единую филогенетическую группу организмов. Данные анализа 16S-рРНК указали на то, чтопрокариоты составляют две основные эволюционные линии: домен Archaea и домен Bacteria. Архебактерии, как уже было сказано, вероятно, одна из самых древних групп живых организмов. Это уникальная группа прокариот, как првило, обитающая в экстремальных местах обитания. Археи осуществляют такие уникальные процессы как метаногенез, бесхлорофилльный фотосинтез. Среди них обнаружены самые термоустойчивые организмы. Среди архебактерий нет патогенных форм, что говорит об их древнем происхождении. В своем метаболизме используют низкомолекулярные соединения, у них не обнаружено гидролаз, поэтому не могут использовать полимеры. Для архей характерны простые циклы развития, у них не обнаружено покоящихся клеток. Морфология. По морфологии не имеют резких отличий от эубактерий. Среди них имеются кокки, палочки, извитые клетки, слабоветвящиеся формы. Грамположительны или грамотрицательные. Особенностью является то, что у них отсутствуют сложные многоклеточные, мицелиальные и трихомные формы. Архебактерии отличаются от эубактерий по ряду признаков: 1. Клеточные стенки архей не содержат пептидогликана, часто состоят из белка, или гетерополисахарида, или псевдомуреина. В связи с этим не чувствительны к антибиотикам, подавляющим синтез пептидогликана, в отличие от эубактерий. 2. Липиды мембран архебактерий представляют собой эфиры глицерина и высокоатомных спиртов (фитанол, бифитанол). 3. Рибосомы по размерам схожи с эубактериальными (имеют константу седиментации 70S), по форме ближе к 80S рибосомам эукариот. Состав рибосомальных РНК архей типично эубактериальный (5S, 16S, 23S), но отличаются по первичной структуре от эубактериальных и эукариотных. 4. Синтез белка у архей не подавляется хлорамфениколом, стрептомицином и другими антибиотиками, подавляющими синтез белка у эубактерий. На рибосомах архей отсутствуют места связывания ингибиторов, специфичных для 70 S рибосом эубактерий. Вместе с тем имеются мета для связывания ингибиторов, специфических в отношении 80S рибосом эукариот. 5. РНК-полимераза архей содержит от 5 до 11 субъединиц. РНК-полимераза эубактерий содержит 4-8 субъединиц. РНК-полимераза эукариот содержит 10-14 субъединиц. РНК-полимераза архей, как и РНК-полимеразы эукариот не чувствительна к антибиотику рифампицину, ингибирующему инициацию, и антибиотику стрептолидигину 1, ингибирующему элонгацию транскрипции. У эубактерий РНК-полимераза чувствительна к этим антибиотикам. 6. Геном архей в виде кольцевой молекулы ДНК. У некоторых архей обнаружены гистоноподобные белки, связанные с ДНК. Особенность генома архей – наличие многократно повторяющихся нуклеотидных последовательностей, а в генах, кодирующих белки, тРНК и рРНК – нитронов, что характерно для организации генетического материала эукариот. Выделяют несколько функциональных групп архебактерий: 1. Метаногены 2. Экстремальные галофилы 3. Экстремальные термофилы и гипертермофилы Метаногены Метанобразующие бактерии осуществляют уникальную в живом мире реакцию синтеза метана. Это облигатные анаэробы. Реакция метаногенеза – единственный источник энергии для этих прокариот. Большинство метаногенов могут расти в газовой смеси Н2 + СО2. При этом они осуществляют следующую реакцию: 4 Н2 + СО2 → СН4 + 2 Н2О Процесс образования метана – это восстановление СО2 до СН4. Донором электронов является Н2, а конечным акцептором электронов СО2 (карбонатное дыхание). СО2 также служит источником углерода. Фиксация СО2происходит по циклическому ацетил-КоА-пути, ключевым промежуточным соединением которог является ацетил-КоА, синтезируемый из двух молекул СО2. Кроме водорода метаногены могут окислять уксусную кислоту, метилированные соединения (напрмер, метанол). По типу обмена метаногены являются хемолитоавторофами (если используют Н2 + СО2) или хемоорганогетеротрофами (если используют уксусную кислоту, метанол). Распространение в природе. Метаногены встречаются в анаэробных местах обитания: В донных отложениях водоемов, в болотах, заболоченных почвах, на рисовых полях, в ЖКТ животных и человека, в рубце жвачных, т.е. там, где идет разложение органического вещества. Метаногены используют продукты брожений (Н2, СО2, уксусная кислота) для образования метана. Практическое значение. Метаногены используются для получения биогаза – метана. Экстремальные галофилы Объединены в порядок Halobacteriales семества Halobacteriacae. Этот порядок включает 6 родов и более 20 видов. Это небольшая группа архей, которые развиваются в средах с концентрацией NaCl от 12-15 до 32 %. Впервые были обнаружены на соленой рыбе. Они поселяются на соленой рыбе и соленых шкурах животных в виде красного налета, вызывая их порчу. Встречаются на солеварнях (в старину в России красную соль считали царской и везли в Москву). В испарительных прудах и лагунах. В засоленных почвах, в гипергалинных водоемах, где концентрация солей – 200 г/л и более, например, в Мертвом море (здесь высокая концентрация не только Na+, но и Mg2+). В щелочных озерах. Являются экстремофилами. При понижении концентрации солей их клетки лизируются. Opt концентрация NaCl – 20-26 %. Галобактерии нуждаются не только в ионах Na+ и Clֿ, но в ионах Mg2+ и К+. Ионы Na+ необходимы для стабилизации клеточной стенки, т.к. у галобактерий преобладают кислые белки, т.е. белки с повышенным содержанием карбоксильных групп (СООН). Ионы Na+ взаимодействуют с отрицательно заряженными молекулами клеточной стенки и придают ей необходимую жесткость. Ионы К+ необходимы для поддержания ионного равновесия вне и внутри клетки. Метаболизм. Галобактерии являются аэробами. Основной способ получения энергии – дыхание. У них имеются дыхательные цепи, полный цикл трикарбоновых кислот. В качестве донора электронов и источника углерода используют органические вещества: аминокислоты, углеводы, органические кислоты. Галобактерии имеют сложные пищевые потребности. Для роста большинства видов в состав сред должны входить дрожжевой экстракт, пептон, гидролизат казеина, набор витаминов. В анаэробных условиях в темноте некоторые виды могут переключаться на нитратное дыхание или брожение, сбраживая аргинин. Фотосинтез – вспомогательный механизм получения энергии, на который переключаются эти бактерии при недостатке в среде О2. У некоторых галобактерий фотосинтез – единственный способ получения энергии. Фотосинтез протекает в бескислородных условиях. В основе бесхлорофилльного фотосинтеза галобактерий лежат светозависимые циклические превращения бактериородопсина. Экстремальные термофилы и гипертермофилы Обитают в потоках гидротерамльной воды и газа в районах вулканической деятельности (Исландия, Камчатка и т.д.), а также в глубоководных морских источниках (гидротермах). Отношение к О2. Аэробы, строгие или фаультативные анаэробы. Метаболизм. Хемоавтотрофы или хемогетеротрофы, могут быть литотрофами (если окисляют Н2) или органотрофами (если окисляют пептон). Являются сульфидогенами, так как участвуют в обмене серы. В анаэробных условиях восстанавливают молекулярную серу до h3S. В аэробных условиях окисляют h3S до молекулярной серы или h3SO4. Представители: Род Sulfolobus – распространены в горячих кислых источниках Температурный диапазон +60 - +90 °С. Гипертермофилы: Наиболее термоустойчивыми на сегодняшний день являются архебактерии-гипретермофилы р. Pyrococcus («неистовые огненные шарики»)и Pyrodictium (огненная сеть). P. furiosus (морская архебактерия, обитает в геотермальных морских осадках) t диапазон 70-103°С, opt t 100°С. Pyrodictium еще более термоустойчива t диапазон 80-110°С, opt t 105 °С (обитает в подводных гидротермах). megaobuchalka.ru Археи — Циклопедия Археи Систематика 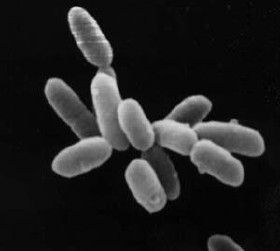 Научная классификация Международное научное название Типы Научная классификация Международное научное название Типы Археи (лат. Archaea от др.-греч. ἀρχαῖος «извечный, древний, первозданный, старый») — одна из групп живых организмов, в которую входят микроскопические одноклеточные прокариоты, которые существенно отличаются рядом физиолого-биохимических признаков от настоящих бактерий. Хотя до сих пор имеется неопределенность в точном филогенезе этих групп, археи, эукариоты и бактерии являются фундаментальными группами в так называемой системе трех доменов. Подобно бактериям, археи — это одноклеточные организмы, не имеющие ядра, и поэтому классифицируются как прокариоты (Prokaryota) — известные также как Monera в таксономии пяти царств. Археи сначала были обнаружено в экстремальных средах, но затем они были найдены во всех типах экосистем. Архебактерии существенно отличаются от других микроорганизмов по составу и последовательности нуклеотидов в рибосомных и транспортных РНК. Археи были идентифицированы в 1977 году Карлом Вёзе (Carl Woese) и Джорджем Фоксом (George Fox), основываясь на их отличиях от других прокариотов (филогенетические деревья, основанные на 16S рРНК). Когда в 1977 году биолог Карл Вёзе изучал генетику микробов, производивших метан, он понял, что они отличаются от всех известных бактерий. Стенка клетки была уникальной — она производила уникальные энзимы и ее генетический ряд не был похож ни на что исследуемое раньше. Очень быстро — в течение часа — стало понятно, что есть еще что-то, третья группа. Это был момент открытия. Карл Вёзе открыл третью форму жизни — группу одноклеточных, которую он назвал археями. Он открыл форму жизни, которая способна существовать где угодно, включая самые невероятные места. Эта группа была сначала названа архебактериями (Archaebacteria) в отличие от эубактерий (Eubacteria), и они рассматривались как царства или подцарства. Вёзе убеждал, что археи являются представителями совсем другой ветви живых существ. Он позже переименовал группы в архей и бактерий, чтобы подчеркнуть это, и аргументировал тем, что вместе с эукариотами они охватывают три группы живых организмов. Биологический термин архея (археи) не следует путать с архейской геологической эрой, также известной как Археозойская эра. Последний термин ссылается на древний период земной истории, когда археи и бактерии были единственными клеточными организмами, жившими на планете. Возможные окаменелости этих микробов были датированы почти 3 800 000 000 лет назад. [править] Археи, эукариоты и бактерии Археи подобны другим прокариотам в большинстве аспектов структуры клетки и метаболизма. Однако, их генетическая транскрипция и трансляция — два центральных процесса в молекулярной биологии — не проявляют типичных для бактерий особенностей, но чрезвычайно подобны этим процессам у эукариотов. Например, система трансляции архей использует эукариотические факторы инициации и элонгации, а их система транскрипции применяет TATA-связываемые белки и TFIIB, как у эукариотов. Гены тРНК и рРНК архей содержат уникальные археальные интроны, которые не похожи ни на эукариотические, ни на бактериальные интроны. Некоторые другие характеристики также выделяют архей. Подобно бактериям и эукариотам, археи имеют основанные на глицерине фосфолипиды. Однако, три особенности липидов архей необычны: - Липиды архей уникальны, так как стереохимия глицерина — обратная той, что найдена у бактерий и эукариот. Это убедительное свидетельство другого биосинтетического пути.
- Большинство бактерий и эукариот имеют мембраны, составленные преимущественно из эфир-липидов глицерина, тогда как археи имеют мембраны, состоящие из эфир-липидов глицерина. Даже когда бактерии имеют эфир-связанные липиды, стереохимия глицерина — бактериальная форма. Эти различия, возможно, являются адаптацией части архей к гипертермофилии. Однако, стоит отметить, что даже мезофиличные археи имеют эфир-связанные липиды.
- Липиды архей основанные на изопреноидной боковой цепочке. Это пятиуглеродная химическая группа, также обычная для каучука и является компонентом некоторых витаминов, распространенна в бактериях и эукариот. Однако, только археи применяют эти вещества в своих клеточных липидах, часто как боковые цепочки C-20 (четыре мономера) или C-40 (восемь мономеров). У некоторых архей изопреноидная боковая цепочка C-40 — фактически достаточно длинная, чтобы пройти сквозь мембрану, формируя монослой вместо клеточной мембраны с группами фосфата глицерина на обоих концах.
Эта адаптация распространенная у чрезвычайно теплолюбивых архей. Хотя они и не уникальны, клеточные стенки архей также необычны. Например, клеточные стенки большинства архей образованны внешними слоями белков или S-слоем. S-слоя распространены у бактерий, они служат единственным компонентом клеточной стенки в некоторых организмах (например, в Planctomyces) или внешним слоем во многих организмах с пептидогликаном. За исключением одной группы метаногенов, археи не имеют пептидогликановой стенки. Однако даже в этом случае, пептидогликаны очень отличаются от разновидности, найденной у бактерий. Археи также имеют жгутики, которые значительно отличаются по составу от очень похожих жгутиков бактерий. Бактериальные жгутики — это измененная система секреции III типа, тогда как жгутики архей напоминают ворсинки IV типа (type IV pili), которые используют sec-зависимую систему секреции, несколько подобную, но все же отличную от системы секреции II типа. В 2016—2017 годах по извлеченным из глубоководных биомов фрагментам ДНК был описан новый надтип — асгардархеи[1]. У них были найдены комплексы многих характерных для эукариот белков, отвечающих за внутренние мембраны, цитоскелет, внутренний транспорт и систему отбраковки белковых дефектов. Выдвинута гипотеза о происхождении эукариот от данной группы архей. Многие археи — экстремофилы. Некоторые живут при очень высоких температурах, часто выше 100° C, как те, которых нашли в гейзерах и черных курильщиках. Других найдены в очень холодных средах или в чрезвычайно соленой, кислой или щелочной воде. Однако, некоторые археи — мезофилы, живут в средах, подобных болотам, сточным водам и почве. Многие метаногенные археи найдены в пищеварительных трактах животных, например, жвачных животных, термитов и людей. Археи не патогенны, и неизвестно, чтобы какие-либо из них вызвали болезнь. Археи разделяются на три группы, в соответствии со средой обитания. Это — галофиты, метаногены и термофилы. Галофиты живут в чрезвычайно соленых окружающих средах (например, многие из них живут в Мертвом море и на юге залива Сан-Франциско, придавая ей яркие цвета: от красного до зеленого). Метаногены живут в анаэробных окружающих средах и производят метан. Их можно найти в отложениях или в кишечниках животных. Термофилы живут в местах с высокими температурами, как, например, горячие источники. Эти группы не обязательно согласуются с молекулярным филогенезом, не полные но не взаимоисключающие. Тем не менее, они — полезная начальная точка для подробного изучения. - The Surprising Archaea: Discovering Another Domain of Life. — Oxford: Oxford University Press, 2000. — ISBN ISBN 0-19-511183-4.
- Giovannoni, S.J. and Stingl, U. (2005). Molecular diversity and ecology of microbial plankton. Nature 437: 343–348.
- Könneke, M., Bernhard, A.E., de la Torre, J.R., Walker, C.B., Waterbury, J.B. and Stahl, D.A. (2005). Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 437: 543–546.
- Lake, J.A. (1988). Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences. Nature 331: 184–186.
- Woese, Carl R.; Fox, George E. (1977). Phylogenetic Structure of the Prokaryotic Domain: The Primary Kingdoms. Proceedings of the National Academy of Sciences of the United States of America 74 (11): 5088-5090.
- Woese, Carl R., Kandler, Otto, Wheelis, Mark L (1990). Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proceedings of the National Academy of Sciences 87 (12): 4576-4579.
cyclowiki.org Архебактерии Архебактерии являются древнейшими прокариотами. Возможно, что они были самыми первыми организмами на Земле.[ ...] Архебактерии играют важную роль в природе.[ ...] Известно около 50 видов архебактерий. Выделяя здесь эту группу организмов в самостоятельное подцарство нельзя не отметить, что многие биологи классифицируют архебактерии в качестве самостоятельного царства (кроме царств растений и животных).[ ...] Классификация архебактерий еще плохо разработана.[ ...] В ДНК некоторых архебактерий отмечается наличие повторяющихся последовательностей азотистых оснований, чего нет у настоящих бактерий. У галофилов обнаружен родопсиноподобный белок, обычно содержащийся в зрительном пурпуре многих позвоночных.[ ...] Прокариоты (бактерии, архебактерии, цианобактерии) — одноклеточные организмы, не имеют ядра. Благодаря такому разнообразному метаболизму бактерии могут существовать в самых различных условиях среды: в воде, воздухе, почве, живых организмах. Велика роль бактерий в образовании нефти, каменного угля, торфа, природного газа, в почвообразовании, в круговоротах азота, фосфора, серы и других элементов в природе. Сапротрофные бактерии участвуют в разложении органических останков растений и животных и в их минерализации до С02, Н20, Н28, 1ЧН3 и других неорганических веществ. Вместе с грибами они являются редуцентами. Клубеньковые бактерии (азотфикси-рующие) образуют симбиоз с бобовыми растениями и участвуют в фиксации атмосферного азота в минеральные соединения, доступные растениям. Сами растения такой способностью не обладают.[ ...] Считают, что первыми возникли архебактерии. Примерно 3,5 млрд лет назад. Затем возникли истинные бактерии, а от них ответвились вирусы, цианобактерии, красные водоросли и зеленые жгутиковые. От последних ответвились водоросли (от которых позднее произошли, в свою очередь, высшие растения) и бесцветные ризофлагелляты, из которых развились диатомовые водоросли, плесневые грибы, губки и простейшие. Часть простейших стала паразитической.[ ...] Перечисленные особенности делают архебактерии по существу промежуточной формой между растениями и животными, что и явилось основанием для некоторых ученых к выделению их в самостоятельное царство.[ ...] Схема синтеза белков, осуществляемого архебактериями, является такой же, как и у настоящих бактерий, однако в тРНК этих организмов нет ни тимина, ни урацила. Есть также отличия и в структуре рРНК.[ ...] В отличие от большинства настоящих бактерий для архебактерии характерен ряд особенностей. Например, плазматическая мембрана архебактерии имеет однослойную структуру, а в пептидогли-кане их клеточных стенок в отличие от клеточных стенок настоящих бактерий отсутствует муреин.[ ...] Уникальностью характеризуются также мембранные липиды архебактерий, ибо они не содержат эфиров глицерина и жирных кислот, но содержат изопреноидные углеводороды, которые обычно встречаются в нефти.[ ...] Самым первым следствием кризиса была гибель подавляющего числа архебактерий в земном океане. Из их остатков сформировались огромные скопления нефти, газа, графитов в отложениях рифея. Вторым важным следствием стало разделение биосферы на две принципиально отличные по жизненным принципам ветви, кстати весьма неравнозначные по объему. Исходная биосфера восстановительного характера уцелела только в донных, преимущественно глубоких зонах, где сформировались локальные резерваты хемобиосферы. На земной поверхности воцарилась оксибиосфера, при этом освободились значительные по объемам экологические ниши. Дальнейший процесс развития новых форм жизни изучен еще слабо, наибольшую обоснованность в его описании имеет теория эндосимбиоза развития по Л. Маргулис. В.А. Зубаков (1997) так описывает ее основные положения: в результате многоэтапных коопераций доядерных органелл с гипотетическим протоядерным организмом уркариотом (по-гречески — творящийся ядерный) возникло новое направление в жизненных формах. Вначале появилась вышеназванная крупная ядерная клетка — эукариот, способная дышать кислородом и генетически активная, совершенствующаяся путем мутаций. Именно генетическая активность резко усилила эволюционный процесс, а главное появилась возможность бесконечного усложнения жизненных форм. И, в частности, кроме всего прочего, жизнь на Земле невозможна без бактерий особых форм, которые определяют завершенность жизненных циклов, а именно выполняют важнейшую функцию деструкции органического материала, синтезируемого эукариотами.[ ...] Ряд ученых выделяет в надцарстве Прокариоты одно царство Дробянки, которое включает три подцарства: Бактерии, Архебактерии и Цианобактерии.[ ...] Однако следует заметить, что многие ученые не разделяют эту точку зрения. Большинство специалистов классифицируют архебактерии в рамках подцарства.[ ...] В этих, уже достаточно суровых, относительно прежних, условиях произошли определенные изменения в органическом мире Земли. Эубактерии или эукариоты, которые к этому времени получили достаточное развитие, представляли собой уже оформленные клеточным ядром образования с отделенной от цитоплазмы ядерной оболочкой. Генетический материал был сформирован в хромосомах. В клетках уже имелись митохондрии, пластиды и другие органоиды. Появление же цианобактерий, использовавших для дыхания азот, привело в конечном счете к постепенному вытеснению архебактерий и эубактерий, что в результате эволюции сделало цианобактерии жизненными монополистами. По В.А. Зубакову (1997), цианобактерии имели очень важную особенность, чрезвычайно существенную для дальнейшего развития органического мира. Этой особенностью было то, что при поглощении азота они выделяли в окружающую водную среду кислород, который в целом был продуктом их жизнедеятельности. Для существовавших же на Земле архебактерий кислород был смертельным ядом. Здесь следует сделать некоторое геологическое отступление.[ ...] В пределах этого надцарства Доядерные организмы выделяют лишь одно царство — царство дробянок (Mychota), которое классифицируют далее на подцарства Архебактерии (Archaeobacteria, или Archaeobacteriobionta), Настоящие бактерии (Bacteria, или Bacteriobionta) и Оксифотобактерии (Oxyphotobacteria, или Oxyphotobacteriobionta).[ ...] Хемоавтотрофы в процессах синтеза органического вещества используют энергию химических связей. К этой группе относятся только прокариоты: бактерии, архебактерии и отчасти синезеленые. Химическая энергия высвобождается в процессах окисления минеральных веществ. Экзотермические окислительные процессы используются нитрифицирующими бактериями (окисляют аммиак до шпротов, а затем до нитратов), железобактериями (окисление закисного железа до окисного), серобактериями (сероводород до сульфатов). Как субстрат для окисления используется также метан, СО и некоторые другие вещества.[ ...] В разных гйзсонах органического мира соотношение форм по Их пищевой сяедааяизащт может быть различным. Чрезвычайно разнообразны по tfttM показателям прокариоты (бактерии, архебактерии, цианобактерии). Они насчитывают относительно небольшое число видов, но представленных большой биомассой, которая обеспечивает высокий уровень Первичной продукции. Показано, например, что в некоторых озерах фотосинтезирующими бактериями создается 3/4 валовой продукций.[ ...] Фотосинтез у прокариот протекает несколько иначе, чем у растений. Бактерии используют для этой функции пигмент бактериохдорин и не выделяют кислород в окружающую среду.[ ...] Кроме вышеописанных организмов-продуцентов в 1887 г. С.Н. Виноградовым были открыты хемосинтезирующие организмы. Эти организмы в процессах синтеза органического вещества используют энергию химических связей. К этой группе продуцентов относят исключительно прокариоты: бактерии, архебактерии и отчасти сине-зеленые. В природе существуют «богатые» потенциальной энергией неорганические соединения. Химическая энергия высвобождается в процессах окисления и некоторых других. Экзотермические (т. е. выделяющие теплоту) окислительные процессы используются азот-фиксирующими (нитрифицирующими) бактериями (окисляют аммиак до нитритов и далее нитратов), железобактериями (окисление закис-лого железа до окисного), серобактериями (сероводород до сульфатов). В частности, последние населяют глубокие океанические впадины, куда не проникает свет, но где в изобилии присутствует сероводород. В этих условиях природа создала уникальные экосистемы, где эти организмы продуцируют органическое вещество в результате хемосинтеза за счет высвобождаемой при расщеплении сероводорода h3S. Как субстрат для окисления используются также метан, оксид углерода и некоторые вещества.[ ...] Максимальный температурный диапазон активной жизни (температура функционирующих клеток) несколько меньше диапазона жидкого состояния воды, а для большинства многоклеточных организмов — от 0 до 50°С. Микроорганизмы-эбулиофилы, обитающие в горячих поверхностных и глубинных источниках, могут сохранять способность к размножению при температуре до 75°С, а некоторые архебактерии нормально существуют при 85—105°.[ ...] Нефтезагрязненные почвы обрабатывают Acinetobacter sp., Alcalgenes sp., Pseudomonas sp., одновременно вносят растворы фосфорных и аммонийных солей. Нефть на поверхности почвы уничтожают бактерии видов Actinomycor elegans и Geotrichum marinum [2, 27]. Использование Actinebacter sp. дает 80%-ный эффект очистки от ароматических соединений по истечении 5 недель [34]. Для деструкции нефти выделены штаммы гало-толерантных и галофильных архебактерий. В условиях высоких температур может быть использована Bacillus thermoleovorans. Бактерия рода Desulfobacterium осуществляет деградацию сали-цилата в условиях сульфатредукции. Деградацию ароматических углеводородов осуществляют некоторые виды Mycobacterium, а также Pseudomonas alcaligenes, которая разлагает и галоуглеводороды [34].[ ...] Предполагают, что жизнь возникла на границе катархея и архея, который начался около 3,5 млрд лет назад и длительность которого составляет около 2 млрд лет. Эта эра характеризовалась широкой вулканической деятельностью. Обнаружение в архейских породах графита и чистого углерода предположительно указывает на существование в этой эре очень простых живых организмов растительной и животной природы, в частности архебактерий и цианобактерий.[ ...] В катархее и архее, на отрезке времени между 4,0—2,8 млрд. лет тому назад, когда сформированная к этому времени атмосфера значительно уплотнилась и насытилась различными парами за счет исключительно активной вулканической деятельности. Это привело к тому, что солнечный свет не достигал поверхности Земли. Высокая плотность атмосферы даже при весьма низкой светимости Солнца (примерно в 4 раза меньше современного) привела к формированию высокого парникового эффекта. Следствием этих факторов стал ровный, очень теплый климат с температурами в диапазоне 200—400° С. Мелкие, пересыщенные солями обширные моря были сосредоточением проявлений жизни. Органический мир был представлен, как пишет В.А. Зубаков, доядерными бессмертными микроорганизмами — прокариотами. Прокариоты были гетеротрофными архебактериями, дышащие метаном и производными серы. Прокариоты не обладали оформленным клеточным ядром и типичным хромосомным аппаратом. Наследственная информация реализовывалась через ДНК.[ ...] ru-ecology.info Настоящие бактерии: чем отличаются представители подцарства С развитием технических средств в классификацию живых организмов постоянно вносятся изменения. Так, в 1977 году Царство дробянок было разделено на 2 части: архебактерии (подцарство древние бактерии) и эубактерии (подцарство настоящие бактерии).  Современная классификация, опираясь на особенности обмена веществ, делит царство дробянок уже на 3 части: - архебактерии;
- настоящие;
- оксифотобактерии или цианобактерии (старое название – сине-зеленые водоросли).
Настоящие бактерии живут на планете более 3 миллиардов лет. Они охватили все экологические ниши – от верхних слоев атмосферы до подводных глубин, живут в полярных областях и рядом с вулканическими образованиями. Они размножаются не только вокруг, но и на поверхности организмов и внутри них. Классификация настоящих бактерий по формеЭубактерии являются одноклеточными организмами, которые по форме клетки подразделяют на три основные группы. КруглыеЭто кокки различной ориентации. В зависимости от особенностей расположения клеток после деления различают: - микрококки – клетки располагаются обособленно;
- диплококки – попарно соединенные клетки;
- стрептококки – объединение круглых бактерий в цепочки;
- сарцины – объединение клеток в плотные пачки;
- стафилококки – конгломераты, строение которых подобно грозди винограда.
ПалочковидныеПалочки после деления могут соединяться попарно – диплобактерии или клебсиеллы, создавать цепочки (стептобактерии) или группироваться в форме креста. Однако большая часть палочковидных бактерий после деления не структурируется и располагается хаотично. Примеры палочковых бактерий: палочка Коха (возбудитель туберкулеза), кишечная палочка, дизентерийная палочка. ИзвитыеСюда относятся спириллы, вибрионы и спирохеты. Пример – бледная спирохета, возбудитель сифилиса. Строение клетки прокариотаНезависимо от формы, все настоящие бактерии, а также архебактерии и цианобактерии имеют характерное строение клетки, содержащее следующие обязательные элементы: - цитоплазму и цитоплазматическую мембрану, окруженную с внешней стороны клеточной оболочкой;
- нуклеоид – невыраженное ядро;
- рибосомы – органеллы клетки прокариотов.
Настоящие бактерии и цианобактерии содержат в составе клеточной стенки муреин, а архебактерии его не имеют, что позволило разделить надцарство Прокариоты на два царства: Эубактерии и Архебактерии. По своему строению архебактерии наиболее схожи с грамположительными настоящими бактериями. Дополнительными деталями можно считать наличие слизистого чехла клетки, а также жгутиков различного количества и расположения, обеспечивающих клетке возможность самостоятельно двигаться. В отсутствие жгутиков клетка способна осуществлять только броуновское движение с потоком жидкости. Жгутики настоящих бактерий характерны только для палочковидных форм. Они являются органом, обеспечивающим прокариотам возможность самостоятельно передвигаться. При этом сам жгутик представляет собой полое белковое тело, сформированное в цитоплазме клетки и выходящее за ее пределы. Характерно, что длина жгутика может значительно превышать размеры самой бактерии. В зависимости от количества и расположения жгутиков, бактерии подразделяют на виды: - монотрихи – единственный жгутик;
- лофотрихи – один пучок;
- амфитрихи – на обоих концах палочки есть один жгутик или пучок;
- перитрихи – жгутики располагаются вокруг клетки равномерно.
Споры как средство выживанияПомимо жгутиков, палочковидные настоящие бактерии отличаются от остальных способностью пережидать неблагоприятные внешние условия, становясь спорами. Спороформирование является сложным процессом, который по продолжительности занимает от 16 до 24 часов. Результатом клеточной перестройки будет бактерия со значительно пониженным количеством воды, а значит, более густой цитоплазмой и особо прочной оболочкой. В форме спор у настоящих бактерий прекращаются обменные процессы, и они не размножаются. Такая защитная форма позволяет настоящим бактериям переносить высокие и низкие температуры, отсутствие влаги, радиоактивное облучение и другие негативные факторы. Будучи спорами, сами бактерии способны сохранять свою жизнеспособность в течение сотен лет. Попадая в благоприятную среду, спора за 4-5 часов «прорастает», превращается в обычную клетку, в которой восстанавливаются строение, обменные процессы и способность размножаться. Ни архебактерии, ни цианобактерии не являются способными к спорообразованию. В зависимости от типа дыхания, спорообразующих аэробов (кислородное дыхание) называют бациллами, а анаэробов (бескислородное) – клостридиями. Классификация по ГрамуНаружная оболочка настоящей бактерии, имеющая достаточно сложное строение, отличается по способности окрашиваться в фиолетовый цвет, что в 1884 г открыл Ганс Кристиан Грам. Эта особенность легла в основу классификации настоящих бактерий как грамположительных (окрашивающихся) и грамотрицательных (неокрашивающихся). Позднее было установлено, что данная способность обусловлена различным составом веществ и организацией клеточной стенки эубактерий. Значение классификации по Граму – упрощение идентификации бактерий, особенно когда они являются патогенными. ПитаниеЭубактерии делят на две группы в зависимости от типа питания: - автотрофные;
- гетеротрофные.
Автотрофные эубактерии способны осуществлять синтез необходимых органических веществ из окружающей неорганики. Их также разделяют на виды по типу энергии, которую микроорганизмы используют при синтезе. Автотрофные настоящие бактерии делят на: - хемосинтезирующие; представители – железобактерии, бесцветные серобактерии и др.;
- фотосинтезирующие; представители – пурпурные и зеленые серобактерии.
Если автотрофные эубактерии получают необходимые вещества, синтезируя их, то гетеротрофные питаются уже имеющимися органическими веществами. Исходя из источника органики, гетеротрофные эубактерии делят на сапрофиты и симбионты (паразиты). Представители сапрофитов перерабатывают органические вещества, источником которых являются неживые организмы. Они ответственны за процессы брожения, когда расщепляются углеродсодержащие вещества, и гниения – когда разлагаются азотсодержащие вещества. Результатом служит выделившаяся энергия, которая используется настоящими бактериями в процессе жизнедеятельности. Значение сапрофитов в круговороте веществ невозможно переоценить, поскольку именно ими разлагаются все природные соединения. Симбиотические эубактерии являются частью здоровой микрофлоры человека, животных и растений. Без них невозможны процессы пищеварения, они участвуют в синтезе некоторых витаминов, расщепляют клетчатку и целлюлозу. РазмножениеНастоящие бактерии размножаются непосредственно прямым бинарным делением – клетка увеличивается в размерах, после достижения критической точки происходит деление на две равновеликие клетки. Архебактерии и цианобактерии являются прокариотами, как и настоящие бактерии, и по способу размножения полностью с ними схожи. Помимо непосредственно прямого деления, для настоящих бактерий характерно несколько способов передачи генетического материала без образования новой клетки. Различают следующие процессы: - конъюгация, когда происходит передача части генетического материала от клетки-донора клетке-реципиенту;
- трансформация – привнесение генетического материала в виде растворимых ДНК в клетку;
- трансдукция – перенос бактериофагами фрагментов генетического материала между различными культурами.
Архебактерии и цианобактерии также способны таким образом разнообразить свой генетический материал. Формирование колонийНастоящие эубактерии размножаются быстро – клетка способна делиться каждые 20 минут. Помимо скорости, большое значение имеет способность ряда настоящих бактерий выделять в окружающую среду активные вещества, негативно действующие на конкурентные виды. Пример – молочнокислые бактерии понижают значение pH окружающей среды, и другие прокариоты не могут расти и размножаться. Или цианобактерии окрашивают воду водоемов – такая среда является токсичной для многих форм жизни. После деления сестринские клетки на какое-то время остаются вместе, образуя цепочки, пакеты или гроздья – так образуются колонии. Для этих конгломератов характерно наличие капсулы из слизи, окружающей всю колонию. Строение колоний, которые образуют цианобактерии, имеют ряд различий от групповых объединений настоящих бактерий. Как пример – рядом расположенные клетки связаны мостиками из цитоплазмы, проходящими клетки насквозь через отверстия в их оболочках. Подобным образом архебактерии, образующие большие колонии, объединяются через длинные трубочки. Кроме того, цианобактерии при объединении в колонии проявляют свойства дифференциации клеток – некоторые отдельные клетки являются носителями дополнительной оболочки. Именно в них осуществляется усвоение азота воздуха. Подобные свойства не свойственны архебактериям или настоящим. Для колонии, состоящей из одного или нескольких типов микроорганизмов не имеют значение соседи – каждая клетка функционирует совершенно автономно. Строение колонии и количество клеток может меняться со временем – это не имеет значения для жизнедеятельности отдельной клетки. probakterii.ru
|















