Гаплокарты: ДНК-генеалогия с высоты птичьего полета. Гаплогруппы самые древние
Гаплогруппы русских: cycyron Оригинал взят у kozzlov56 в Гаплогруппы русскихГаплогруппы русских Природой, генетический код у всех людей устроен таким образом, что у каждого имеются 23 пары хромосом, в которых хранится вся наследственная информация, доставшаяся от обоих родителей. Формирование хромосом происходит в момент мейоза, когда в процессе кроссинговера, каждая случайно берет примерно половину от материнской хромосомы и половину от отцовской, какие конкретно гены достанутся от мамы, а какие от папы - не известно, все решает случай.В этой лотереи не участвует только одна мужская хромосома – Y, она целиком передается от отца к сыну как эстафетная палочка. Уточню, что у женщин данной Y-хромосомы нет совсем.В каждом последующем поколении в определенных участках Y-хромосомы, называемых локусами, происходят мутации, которые будут передаваться всем последующим поколениям по мужскому роду. Именно благодаря этим мутациям стало возможным реконструировать рода. На Y хромосоме всего примерно 400 локусов, но для сравнительного анализа гаплотипов и реконструкции родов используется только около сотни.В так называемых локусах, или их еще называют STR-маркеры, бывает от 7 до 42 тандемных повторов, общая картина которых является уникальной для каждого человека. Через определенное количество поколений происходят мутации и количество тандемных повторов изменяется в большую или меньшую сторону и таким образом на общем древе будет видно, что чем больше мутаций, тем древнее общий предок для группы гаплотипов. Сами гаплогруппы не несут генетической информации, т.к. генетическая информация находиться в аутосомах- первых 22 пар хромосом. Посмотреть распространение генетических компонент в Европе можно здесь. Гаплогруппы являются всего лишь метками давно минувших дней, на заре формирования современных народов. Какие гаплогруппы у русских наиболее часто встречаются?Народы Кол-во,человек R1a1,% R1b1,% I1,% I2,% N1c1,% E1b1b1,% J2,% G2a,% | Восточные, западные и южные славяне. | | Русские (север) | 395 | 34 | 6 | 10 | 8 | 35 | 2 | 1 | 1 | | Русские (центр) | 388 | 52 | 8 | 5 | 10 | 16 | 4 | 1 | 1 | | Русские (юг) | 424 | 50 | 4 | 4 | 16 | 10 | 5 | 4 | 3 | | Русские (все великороссы) | 1207 | 47 | 7 | 5 | 12 | 20 | 4 | 3 | 2 | | Белорусы | 574 | 52 | 10 | 3 | 16 | 10 | 3 | 2 | 2 | | Украинцы | 93 | 54 | 2 | 5 | 16 | 8 | 8 | 6 | 3 | | Русские (вместе с украинцами и белорусами) | 1874 | 48 | 7 | 4 | 13 | 16 | 4 | 3 | 3 | | Поляки | 233 | 56 | 16 | 7 | 10 | 8 | 4 | 3 | 2 | | Словаки | 70 | 47 | 17 | 6 | 11 | 3 | 9 | 4 | 1 | | Чехи | 53 | 38 | 19 | 11 | 12 | 3 | 8 | 6 | 5 | | Словенцы | 70 | 37 | 21 | 12 | 20 | 0 | 7 | 3 | 2 | | Хорваты | 108 | 24 | 10 | 6 | 39 | 1 | 10 | 6 | 2 | | Сербы | 113 | 16 | 11 | 6 | 29 | 1 | 20 | 7 | 1 | | Болгары | 89 | 15 | 11 | 5 | 20 | 0 | 21 | 11 | 5 | | Балты, финны, германцы, греки и др. | | Литовцы | 164 | 34 | 5 | 5 | 5 | 44 | 1 | 0 | 0 | | Латыши | 113 | 39 | 10 | 4 | 3 | 42 | 0 | 0 | 0 | | Финны (восток) | 306 | 6 | 3 | 19 | 0 | 71 | 0 | 0 | 0 | | Финны (запад) | 230 | 9 | 5 | 40 | 0 | 41 | 0 | 0 | 0 | | Шведы | 160 | 16 | 24 | 36 | 3 | 11 | 3 | 3 | 1 | | Немцы | 98 | 8 | 48 | 25 | 0 | 1 | 5 | 4 | 3 | | Немцы (баварцы) | 80 | 15 | 48 | 16 | 4 | 0 | 8 | 6 | 5 | | Англичане | 172 | 5 | 67 | 14 | 6 | 0.1 | 3 | 3 | 1 | | Ирландцы | 257 | 1 | 81 | 6 | 5 | 0 | 2 | 1 | 1 | | Итальянцы | 99 | 2 | 44 | 3 | 4 | 0 | 13 | 18 | 8 | | Румыны | 45 | 20 | 18 | 2 | 18 | 0 | 7 | 13 | 7 | | Осетины | 359 | 1 | 7 | 0 | 0 | 1 | 16 | 67 | | Армяне | 112 | 2 | 26 | 0 | 4 | 0 | 6 | 20 | 10 | | Греки | 116 | 4 | 14 | 3 | 10 | 0 | 21 | 23 | 5 | | Турки | 103 | 7 | 17 | 1 | 5 | 4 | 10 | 24 | 12 |
нно обращают на себя внимание 4 наиболее распространенных среди русских гаплогруппы:R1a1 47.0%, N1c1 20.0%, I2 10.6%, I1 6.2%Говоря простыми словами: генетический состав русских по прямым мужским линиям Y-хромосомы выглядит следующим образом:восточно-европейцы - 47%балтийцы - 20%И две гаплогруппы исконных европейцев со времен палеолитаскандинавы - 6%балканцы - 11%Названия условны и даны в соответствии с территориальными максимумами европейских субкладов для гаплогрупп R1a1, N1c1, I1 и I2. Принципиальным моментом является то, что потомков монголов после двухсотлетнего татаро-монгольского ига не осталось. Либо осталось, но очень малое количество прямых генетических наследников от таких связей. Данными словами я ни сколько не хочу подвергать сомнению исторические источники о монголах на Руси, а только обратить внимание на предполагавшееся генетическое влияние со стороны монголо-татар на русских - его нет, либо оно незначительно. Кстати сказать, в геноме булгарских татар также большое количество носителей гапрогруппы R1a1 (около 30%) и N1c1 (около 20%), но они по большей степени не европейского происхождения. Еще один важный момент, южные русские в пределах погрешности не отличаются от украинцев, a северные русские имея одной из преобладающих ту же гаплогруппу R1a1, также имеют более высокий процент гаплогруппы N1c1. Но % N1c1 гаплотипов в среднем 20% у русских. Николай 2 Первым известным предком Ольденбургского великогерцогского Дома был, упоминаемый в летописях за 1091 год, Эгильмар, граф Леригау (ум. 1108 ).Николай II оказался носителем гаплогруппы R1b1a2 - представителем западноевропейской линии, от династии Гольштейн-Готторпов. Для данной германской династии характерен терминальный снип U106, имеющий наибольшее распространение в северо-западной Европе в местах расселения германских племен. Это не совсем типичный для русского народа ДНК маркер, но его наличие среди русских могло быть также связано с ранними контактами германцев и славян. РюриковичиВладимир Мономах и его потомки, именуемые «Мономашичами» принадлежат к гаплогруппе N1c1-L550, которая имеет наибольшее распространение в Южно-Балтийском регионе (субклад L1025) и в Фенноскандии (субклады Y7795, Y9454, Y17113, Y17415, Y4338). Для династии Рюриковичей характерен терминальный снип Y10931.Некоторые из тех, кого историки называют Ольговичами (названы в честь Олега Святославовича - главного соперника Владимира Мономаха в феодальной борьбе - и, как уверяют все источники, его двоюродного брата) к Рюриковичам из рода Мономашичей не имеют родственного отношения (по прямой мужской линии).Подробнее >>> Русские, славяне, индо-европейцы и гаплогруппы R1a, R1b, N1c, I1 и I2В глубокой древности, около 8-9 тысячелетий назад была лингвистическая группа, положившая начало индо-европейской семье языков (на начальном этапе вероятнее всего это гаплогруппы R1a и R1b). Индоевропейская семья включает в себя такие лингвистические группы, как индо-иранцы (Южная Азия), славяне и балты (Восточная Европа), кельты (Западная Европа), германцы (Центральная, Северная Европа). Возможно, у них были и общие генетические предки, которые около 7 тыс. лет назад вследствие миграций оказались в разных частях Евразии, часть ушла на юг и восток (R1a-Z93), положив начало индо-иранским народам и языкам (во многом приняв участие в этногенезе тюркских народов), а часть осталась на территории Европы и положила начало образованию многих европейских народов (R1b-L51), включая славян и русских в частности (R1a-Z283, R1b-L51). На разных этапах формирования уже в древности были пересечения миграционных потоков, что послужило причиной наличия у всех европейских этносов большого количества гаплогрупп.Славянские языки выделились из некогда единой группы балто-славянских языков (предположительно археологическая культура поздней шнуровой керамики). По расчетам лингвиста Старостина это произошло примерно 3,3 тысячелетия назад. Период от V века до н.э. по IV-V век н.э. можно считать условно праславянским, т.к. балты и славяне уже разделились, но самих славян еще не было, они появятся несколько позже, в 4-6 веках н.э. На начальном этапе формирования славян вероятно около 80% составляли гаплогруппы R1a-Z280 и I2a-M423. На начальном этапе формирования балтов вероятно около 80% составляли гаплогруппы N1c-L1025 и R1a-Z280. Влияние и пересечение миграций балтов и славян было с самого начала, потому во многом это деление условно, и в целом отражает только основной тренд, без подробностей. Иранские языки относятся к индоевропейским, и датировка их следующая – древнейшие, от 2-го тысячелетия до н.э. по IV век до н.э., средний – от IV века до н.э. по IX век н.э., и новый – с IX века н.э. по настоящее время. То есть древнейшие иранские языки появляются уже после ухода части племен говоривших на индо-европейских языках из Средней Азии в Индию и Иран. Основными гаплогруппами у них вероятно были R1a-Z93, J2a, G2a3. Западно-иранская группа языков появилась уже позже, примерно в V веке до н.э. Таким образом индо-арии, кельты, германцы и славяне в академической науке стали индоевропейцами, этот термин наиболее адекватный для такой такой обширной и разнообразной группы. Это совершенно корректно. В генетическом аспекте бросается в глаза неоднородность индо-европейцев как по Y-гаплогруппам, так и по аутосомам. Для индо-иранцев характерно в большей степени переднеазиатское генетическое влияние БМАК. Согласно индийским ведам, именно индо- арии пришли в Индию (в Южную Азию) с севера (из Средней Азии), и это их гимны и сказания легли в основу индийских вед. И, продолжая дальше коснемся лингвистики, ведь это русский язык (и родственные ему балтийские языки, например, литовский в составе когда-то существовавшей балто-славянской языковой общности) относительно близок к санскриту наравне с кельтскими, германскими и другими языками большой индо-европейской семьи. Но в генетическом плане индо-арии были уже в большей степени переднеазиатами, по мере приближения к Индии усиливалось и веддоидное влияние. Итак, стало ясно, что гаплогруппа R1a в ДНК-генеалогии – это общая гаплогруппа для части славян, части тюрков и части индо-ариев (т.к. естественно в их среде были представители и других гаплогрупп), часть гаплогруппы R1a1 при миграциях по Русской равнине вошли в состав финно-угорских народов, например мордвы (эрзя и мокша). Часть племен (для гаплогруппы R1a1 это субклад Z93) при миграциях принесли этот индо-европейский язык в Индию и Иран примерно 3500 лет назад, то есть в середине II-го тыс. до нашей эры. В Индии он трудами великого Панини был произведен в санскрит в середине I-го тыс. до н.э., а в Персии-Иране арийские языки стали основой группы иранских языков, древнейшие из которых датируются II-м тысячелетием до н.э. Эти данные подтверждаются: ДНК-генеалогия и лингвистика здесь коррелируют между собой. Обширная часть гаплогруппы R1a1-Z93 еще в древности влились в тюркские этносы и маркируют сегодня во многом миграции тюрков, что и неудивительно ввиду древности гаплогруппы R1a1, в то время как представители гаплогруппы R1a1-Z280 входили в финно-угорские племена, но при поселении славянских колонистов многие из них были ассимилированы славянами, однако даже сейчас у многих народов например у эрзя по-прежнему доминирующей гаплогруппой является R1a1-Z280.Все эти новые данные смогла нам предоставить ДНК-генеалогия, в частности, примерные даты миграций носителей гаплогрупп на территории современных Русской Равнины и Средней Азии в доисторическое время.Так ученые всем славянам, кельтам, германцам и т.д. дали имя индоевропейцев, что соответствует действительности с точки зрения лингвистики.Откуда появились эти индоевропейцы? На самом деле индо-европейские языки были задолго до миграций в Индию и Иран, по всей Русской равнине и до Балкан на юге, а до Пиреней на западе. В дальнейшем язык был распространен в Южную Азию - и в Иран и в Индию. Но в генетическом плане корреляций намного меньше.«Единственно оправданным и принятым в настоящее время в науке является применение термина «арии» лишь по отношению к племенам и народам, говорившим на индо-иранских языках». И далее – «В языкознании арийскими называются индо-иранские языки». Так в какую сторону шел индо-европейский поток – на запад, в Европу, или наоборот, на восток? По некоторым оценкам индоевропейской языковой семье около 8500 лет. Прародина индо-европейцев до сих пор не определена, но по одной из версий это могло быть Причерноморье- южное или северное. В Индию, как мы уже знаем индо-арийский язык был привнесен около 3500 лет назад предположительно с территории Средней Азии, а сами арии были группой с различными генетическими Y-линиями, как R1a1-L657, G2a, J2a, J2b. Гаплогруппа R1a1 в Западной и Южной ЕвропеАнализ 67-маркерных гаплотипов гаплогруппы R1a1 из всех стран Европы позволил определить примерный путь миграции предков R1a1 в направлении Западной Европы. И расчеты показали, что почти по всей Европе, от Исландии на севере до Греции на юге, общий предок у гаплогруппы R1a1 был один приблизительно 7500 лет назад! Иначе говоря, потомки как эстафету передавали свои гаплотипы своим же потомкам из поколения в поколение, расходясь в процессе миграций из одного и того же исторического места - которым оказались предположительно Балканы или Причерноморская низменность. И не просто Балканы, а Сербия, Босния, Македония, а также Белоруссия, Украина, Россия. Это – ареал самых древних гаплотипов гаплогруппы R1a1. И время жизни первопредка, на которое указывают самые древние, самые мутированные гаплотипы – это 7,5 тысяч лет назад. В те времена еще не было ни славян, ни германцев, ни кельтов.ДНК-генеалогия вполне определенно указывает, что на протяжении многих тысячелетий дальние предки современных людей жили в характерных ареалах, никуда особо не передвигаясь. Если и передвигались – следов тех активистов в гаплотипах наших современниках пока не найдено. Возможно, и не осталось. Но примерно 6 тысяч лет назад начались массовые миграции, которые в дальнейшем оставят свой след в истории всей Европы - это и миграции тюрков, уральцев, и собственно индо-европейцев, начались они в связи с переходом к новым формам хозяйствования и необходимостью освоения новых территорий, эти миграции можно назвать постнеолитическими.Ряд археологических культур в Европе до сих пор не обрел лингвистическую принадлежность. Много было палео-европейских языков, от которых в Европе остался только язык басков и некоторая часть лексики в языках северной Европы. Одной из таких является Среднестоговская культура. Их общий предок, по местным гаплотипам, жил 6 тысяч лет назад, и принадлежал возможно к гаплогруппе R1a1 и R1b.В дальнейшем пошли расходиться миграционные потоки, где возможно также основной была гаплогруппа R1a1 во все стороны, с Балкан, с Причерноморья - Ямная археологическая культура, Хвалынская археологическая культура) и Буковины (Трипольская культура). Практически во все стороны – 6-5 тысяч лет назад, третье-четвертое тысячелетие до нашей эры. Где была до этого момента гаплогруппа R1a1 сказать сложно, но предполагается что в Европу она попала из Азии. Германия – базовый 67-маркерный гаплотип имеет свои характерные мутации и разделение с восточными славянами произошло приблизительно 6 тыс. лет назад. Субклад M458.Сейчас его обладателей (уже с мутациями) в Германии в среднем 14% (R1a1-M458), но в некоторых районах достигает трети. Большинство остального населения Германии имеют «скандинавскую» гаплогруппу I1 (28%) и «западноевропейскую» R1b1a2 (39%).Норвегия – уже другой гаплотип, предок на территории современной Норвегии жил 4500 лет назад. В Норвегии доля R1a1-Z284 сейчас в среднем – от 18 до 25% населения. В основном – скандинавская I1 (41%) и западноевропейская R1b1a2 (28%) гаплогруппы. У норвежцев свой субклад этой древней гаплогруппы R1a1-Z284. Поскольку у всех прочих европейцев гаплогруппы R1a1 имеются различные субклады, то не буду это больше и упоминать. Буду только указывать, когда первопредок гаплогруппы R1a1) жил и к какому субкладу относился на каждой конкретной территории.Швеция – 4500 лет назад. Видно, что для освоения северных территорий – Норвегии, Швеции – понадобилось время, несколько веков, 500-600 лет. Всего среди современных шведов 17% потомков гаплогруппы R1a1-Z284. В основном – скандинавская I1 (48%), западноевропейская R1b (22%) и балтийская N1c1 (11%) гаплогруппы. Перейдем к Атлантике, на британские острова. Здесь – целая группа территорий, на которых издавна живут потомки гаплогруппы R1a1-L664. Они численно отнюдь не доминируют по сравнению с другой гаплогруппой R1b, типичным представителем которой являются кельты, и которые пришли туда 3500-4000 лет назад. Но и их, потомков гаплогруппы R1a, на островах не так мало. Они выделяются наличием уникальной точечной мутацией L664. В Англии общий предок современных носителей R1a1 жил 4500 лет назад, как и в Германии. Но в Англии и вообще на британских островах потомков R1a1 относительно мало, от 2% до 9% по всем островам. Там полностью доминируют западноевропейская гаплогруппа R1b (71%) и скандинавская гаплогруппа I1 (16%).В Ирландии – 5200 лет назад. Ирландские гаплотипы - самые древние в Западной Европе и на Британских островах для гаплогруппы R1a1. причина - уникальный субклад L664. Очень вероятно, что заселение было очень ранним, а также возможно выжили древние ирландские R1a1 удачнее материковых. Но сейчас в Ирландии представителей гаплогруппы R1a1 немного, не более 2-4% населения. Там более, чем три четверти западноевропейской гаплогруппы R1b1a2.На освоение северной, холодной и горной Шотландии понадобилось время. Общий предок здешнего субклада гаплогруппы R1a1 жил 4300 лет назад. В Шотландии потомки R1a1 по численности снижаются с севера на юг. На севере, на Шетландских островах, их 27%, и эта численность падает до 2-5% на юге страны. В среднем, по всей стране, их около 6%. Остальные – от двух третей до трех четвертей – имеют западноевропейскую гаплогруппу R1b. Центральная и Восточная ЕвропаПольша, общий предок R1a1 жил около 5000 лет назад (в основном это субклад R1a1-M458 и Z280). У русско-украинских – 4500 лет назад, что практически совпадает в пределах точности расчетов. Да и даже если четыре поколения – это не разница для таких сроков. В современной Польше гаплогруппы R1a1 в среднем 56%, и в некоторых районах до 62%. Остальные имеют в основном западноевропейскую гаплогруппу R1b (12%), скандинавскую гаплогруппу I1 (17%) и балтийскую гаплогруппу N1c1 (8%).В Чехии и Словакии общий праславянский предок жил 4200 лет назад. Всего ненамного меньше, чем у русских и украинцев. То есть речь идет о расселении на территориях современных Польши, Чехии, Словакии, Украины, Белоруссии, России – все в пределах буквально нескольких поколений, но четыре с лишним тысячи лет назад. В археологии такая точность датировок совершенно немыслима. В Чехии и Словакии потомков гаплогруппы R1a1 около 40%. У остальных в основном западноевропейская R1b (22-28%), скандинавская I1 и балканская гаплогруппа I2a (в совокупности 18%) На территории современной Венгрии общий предок R1a1 жил 5000 лет назад. Там сейчас до четверти потомков гаплогруппы R1a1. Остальные имеют в основном западноевропейскую гаплогруппу R1b (20%) и совокупную скандинавскую I1 и балканскую I2 (суммарно 26%) гаплогруппы. Учитывая, что венгры говорят на языке финно-угорской группы языков, наиболее распространенной гаплогруппой у которых является N1c1 в древних венгерских богатых захоронениях мадьяр в основном находят останки мужчин с гаплогруппой N1c1, которые и были первыми вождями племен, участвовавших в становлении империи. В Литве и Латвии общий предок реконструируется на глубину 4800 лет. В основном там сегодня субклад Z92, Z280 и M458. Наиболее распространена среди литовцев балтийская гаплогруппа N1c1, достигая 47%. Вообще же для Литвы и Латвии характерен Южно-Балтийский субклад L1025 гаплогруппы N1c1. В общем, ситуация ясна. Добавлю только, что по европейским странам - Исландии, Нидерландов, Дании, Швейцарии, Бельгии, Литвы, Франции, Италии, Румынии, Албании, Монтенегро, Словении, Хорватии, Испании, Греции, Болгарии, Молдавии – общий предок жил 5000-5500 лет назад, точнее установить невозможно. Это – общий предок гаплогруппы R1a по всем перечисленным странам. Общеевропейский предок, так сказать, не считая показанного выше балканского региона, возможной прародины, индоевропейцев около 7500 лет назад. Доля носителей гаплогруппы R1a1 в следующих странах варьируется, от 4% в Голландии и Италии (до 19% в Венеции и Калабрии), 10% в Албании, 8-11% в Греции (до 25% в Салониках), 12-15% в Болгарии и Герцоговине, 14-17% в Дании и Сербии, 15-25% в Боснии, Македонии и Швейцарии, 20% в Румынии и Венгрии, 23% в Исландии, 22-39% в Молдавии, 29-34% в Хорватии, 30-37% в Словении (16% в целом по Балканам), ну и заодно – 32-37% в Эстонии, 34-38% в Литве, 41% в Латвии, 40% в Белоруссии, 45-54% на Украине. В России восточно-европейской гаплогруппы R1a, как я уже упоминал, в среднем 47%, за счет высокой доли балтийской гаплогруппы N1c1 на севере и северо-западе России, но на юге и в центре России доля разных субкладов гаплогруппы R1a достигает 55%. Тюрки и гаплогруппа R1a1Гаплотипы предков везде разные, для разных регионов характерны свои субклады. У народов Алтая и других тюрков также высокие проценты гаплогруппы R1a1, у башкир достигает 40% субклад Z2123. Это дочерняя линия от Z93 и её можно назвать типично тюркской и не имеющей отношения к миграциям индо-иранцев.Сегодня большое количество гаплогруппы R1a1 находиться в Саяно-Алтайском регионе, среди тюркского населения Средней Азии. У киргизов достигая 63%. Ни русскими ни иранцами их не назовешь. Получается назвать всю гаплогруппу R1a1 единым названием - грубое утрирование, по меньшей мере, а по большей - невежество. Гаплогруппы - это не этносы, на них не записана лингвистическая и этническая принадлежность носителя. К генам тоже прямого отношения гаплогруппы не имеют. Для тюрков в основном характерны различные субклады Z93, но в Поволжье встречаются и R1a1-Z280, возможно перешедшие к поволжским тюркам от поволжских финнов. Гаплогруппа R1a1-Z93 также в умеренной частоте характерна для арабов, для левитов - подгруппы ашкеназских евреев (у последних был подтвержден субклад CTS6). Данная линия уже на самых ранних этапах приняла участие в этногенезе этих народов.Территория первоначального распространения гаплогруппы R1a1 в Европе - это, вероятно, территория Восточной Европы и возможно Причерноморская низменность. До этого вероятно в Азии, возможно в Южной Азии или Северном Китае. Кавказские гаплотипы R1a1Армения. Возраст общего предка гаплогруппы R1a1 – 6500 лет назад. В основном тоже субклад R1a1-Z93, хотя есть и R1a1-Z282.Малая Азия, Анатолийский полуостров. Исторический перекресток на путях между Ближним Востоком, Европой и Азией. Это был первый или второй кандидат для «индоевропейской прародины». Однако общий предок гаплогруппы R1a1 жил там около 6500 лет назад. Ясно, что этой прародины, судя по гаплотипам, в Анатолии быть практически может, либо первоначальными индо-европейцами были носители гаплогруппы R1b. Но высока вероятность низкой представленности в общей базе гаплотипов индивидов из Турции.Итак, и армяне, и анатолийцы – у всех предок или тот же самый, или предки очень близки по времени, в пределах нескольких поколений - это субклад Z93 и Z282*. Следует отметить, что 4500 лет до общего предка гаплогруппы R1a1-Z93 в Анатолии хорошо согласуется со временем появления хеттов в Малой Азии в последней четверти III тысячелетия до н.э, хотя многие линии R1a1-Z93 там могли появиться после миграций тюркских народов на полуостров уже в нашей эре. cycyron.livejournal.com Гаплогруппы в этноопределении это Миф - 20 Января 2014 - MerjamaaВ различного рода псевдоисторических рассуждениях, касающихся исторического происхождения россиян исключительно из славян, теперь модно вплетать и "гаплогруппы". С легкой руки некоего "ученого" (совсем не биолога) Клесова, проживающего (как и "академик" Левашов) в США (отсюда до смешного знакомый приемчик - аппеляция к авторитету "западной науки" - "американские ученые доказали что русским 4500 лет!") этот вполне научный метод определения отцовства и династичности стал использоваться для построений, для которых он совсем не предназначен. В итоге многие посты содержат чудовищно искаженные результаты его использования. Целью этого поста я ставил прояснить очевидные моменты, дабы многочисленные, приводимые авторами к месту и не к месту, ссылки на то что "гаплогруппы доказали" не были воспринимаемы некритично при чтении некоторых текстов. Позволю себе прояснить некоторые общие места в виде популярно данных ответов на вопросы. Вопрос 1 Может ли конкретная гаплогруппа указывать на определенную национальность? Иначе говоря существует ли русская или немецкая (без кавычек) гаплогруппа? Ответ: Нет. Почему? Потому, что любая ныне существующая нация этнос состоит из людей как минимум с двумя-тремя Y-хромосомными гаплогруппами. Иными словами вы можете быть русским с гаплогруппой и I1, и R1a1, и R1b, N3, I2, J2 и даже E. То же касается и остальных народов. например среди евреев-ашкенази - около 12% носителей якобы "маркерной" для русских гаплогруппы R1a1 - причем подчеркивается, что не у «полукровок» и "гоев", а у самых настоящих, типических представителей своего этноса. И так далее. Таким образом, соотнесение определенной гаплогруппы с современными национальностями и отдельными представителями этносов может носить лишь статистический, относительный характер. "Измерять черепа" по гаплогруппам ну никак не получится - вполне вероятно, что еврея вы запишете в русские, а русского - в евреи. О том, можно ли определять народы как «арийцев» только по одному критерию - по наличию статистически маркерной гаплогруппы, говорит наличие R1a у поляков - 56,4%, и… киргизов - аж 63,5%. А волжские татары по набору гаплогрупп вообще ничем не отличаются от русских. Надеюсь, что выводы сделаны? Иначе говоря, никакой этнической общности, объединяющей людей из условно «арийской» гаплогруппы R1a1, не выйдет. В то же время и никакой объективной разницы между живущими в одной области классическими русскими, но с условно «финской» гаплогруппой N, и теми же классическими русскими, носителями «арийской» гаплогруппы R1a, не существует. Весь остальной генофонд (и во многом более более значимый, чем гапломаркер) у потомков «предковых мужчин-финнов» и «предковых мужчин-ариев» давным давно перемешался. Вопрос 2. О чем говорят гаплогруппы на самом деле и о чем говорить не могут? Ответ: Как вытекает из ответа 1 и как правильно отметил автор еще одного обстоятельного поста, посвященного гаплогруппам: "Те участки Y-хромосомы, которые служат маркерами для выделения гаплогрупп, сами по себе ничего не кодируют и никакого биологического смысла не имеют. Это маркеры в чистом виде. Их можно сравнить с оранжевым и зеленым светодиодами в фильме «Кин-дза-дза», которые использовались для идентификации чатлан и пацаков, причем никакой другой разницы, кроме цвета лампочки, между этими «расами» не было". По гаплогруппам можно судить - статистически (!!! это важно - именно статистически, т.е. с неизбежными погрешностями, но все же можно) о разнице, несходстве в происхождении тех или иных больших этнолингвистических массивов и степени близости их контактов (половых причем). Например, все тюркские народы являются носителями специфических гаплогрупп. Даже если у части тюрков нет или очень мало их "маркерных" и отстутствующих у славян C, O и Q, то и реже встречаемые у тюрков D, R1b1b1 или N1b, все равно статистически отсутствуют у русских, украинцев и белоруссов. Можно сделать вывод, т.о.,что контакты с тюрками на протяжении истории у восточных славян - статистически незначительны, а утверждение о том что "мы - азиаты" - неверно. Такие выводы вполне научны. Можно утверждать также, что типичные «монголоидные» гаплогруппы C и Q у русских вообще не встречаются, а раз так, контакты населения Руси с монголами в период ига были минимальными. Об этом, впрочем, ясно свидетельствуют и все исторические источники - спустя 20 лет после 1240 года все монгольские военные формирования были выведены с территории Руси и русские князья возили дань, собираемую на Руси в Орду сами. Таким образом,скорее всего среднестатитстический русский крестьянин конца 13 и 14-15 вв никогда в своей жизни не видел ни одного живого "монгола" (ордынца). И это - никакое не "открытие", - гаплогруппы лишь подтверждают то, что нам известно и так (другое дело, что в "популярной" антиисторической литературе, в т.ч. и в учебниках, содержание которых ныне практически не контролируют ученые, об этом мало пишут). Далее - из ответа 1 ясно, что если представители какого-то этноса относятся к нескольким гаплогруппам, то это НЕ ЗНАЧИТ, что данный этнос представляет собой некое механическое соединение популяций с разным генофондом. Весь остальной генофонд, кроме Y-хромосом, у них будет перемешан без практической возможности различия. 16% гаплогруппы N у русских в регионах севернее Москвы не означает что 16% из них якобы "больше финны, чем славяне", - и по причине, указанной выше (перемешивание до степени неузнаваемости всего остального, гораздо более важного, генофонда) и по причинам статистическим - т.к. у населения Финляндии - лишь 60% населения имеют эту гаплогруппу, а среди эстонцев таких вообще - 40%. Я опять процитирую - "Особенность Y хромосомы в том, что она передается от отца к сыну практически в неизменном виде и не испытывает «смешения» и «разбавления» материнской наследственностью. Это позволяет использовать ее как математически точный инструмент для определения происхождения по отцовской линии. Если термин «династия» имеет какой-либо биологический смысл, то это именно наследование Y-хромосомы. Y-хромосомная гаплогруппа определяет совокупность мужчин, объединенных наличием такого маркера, т.е. происходящих от общего предка-патриарха, у которого много тысяч лет назад случилась специфическая мутация Y-хромосомы." Современная наука склоняется к тому, что эти мутации могли произойти в промежутке от 50 до 10 тыс лет назад, а креационистская версия, кажущаяся мне более правдивой, говорит о совершеннейшей неопределенности результатов использования методов "биологических часов" (они дают совершенно разные значения ) и говорит о вероятной практической одномоментности появления этих мутаций, связываемых с легендой о Вавилонской башне и рассеянием народов. Правы ли те или другие, для нас, ныне живущих, в принципе не очень важно. Важно то, что Y - хромосома не определяет генофонд. Не является носителем некоей "этнически определенной" информации в геноме. Из более чем 20000 генов человеческого генома, в Y-хромосому входит всего около 100 штук. Они кодируют в основном устройство и функционирование мужских половых органов. Никакой другой информации там нет. Черты лица, цвет кожи, особенности психики и мышления прописаны в других хромосомах, которые при наследовании проходят через рекомбинацию (отцовские и материнские участки хромосом перемешиваются случайным образом). Носителем "арийской хромосомы" спустя сотни поколений может быть типичный негр (если. скажем, мама-негритянка залетела в 16 веке от португальца). И наоборот. Таким образом. люди которые говорят об "арийских", "семитских", "русских", "финских", "кельтских" гаплогруппах не в их условно-маркерном значении, привязанном к СОВРЕМЕННОСТИ, а в буквально понимаемом, да еще и переносят эти определения в далекое прошлое - заблуждаются и путают других. Фраза "4500 лет назад в Воронежской области нашли скелет, по гаплогруппе определяемый как русский, русские начали существовать 4500 лет назад" - не имеет никакого смысла. Русские имеют несколько гаплогрупп, - раз. Скелет не представляет собой статистически значимую выборку - это два (может это тот самый еврей-ашкенази из 12% носителей "арийской" гаплогруппы R1a1? или киргиз?). Три: с какой стати сделан вывод что мутация возникла (и имела продолжение) именно 4500 лет назад? Разве были исследованы скелеты его папы и дедушки, а также скелеты потомства? Почему она не возникла 10 тыс лет назад? И так далее. Вопрос 3: является ли R1a1 - русской гаплогруппой, "безусловным эническим маркером"? Правда ли, что русские по своей чистоте (т.е. частоте встречаемости этой гаплогруппы) - самые устойчивые и чистые среди европейцев? Ответ: Нет. И неправда. Устойчивость русских как народа, его этническая уникальность не определяется гаплогруппами, и гаплогруппой R1a1 - тоже. Смотрим данные: если делать усредненную выборку из нескольких разных исследований с подгонкой под 100%. то (помним, что гаплогруппы имеют название по СОВРЕМЕННОЙ дистрибуции среди народов): У 47% русских представлена «арийская» гаплогруппа R1a1, которую они унаследовали от праславян и других автохтонов Восточной Европы. У 16% русских - условно «финская» гаплогруппа N3. У 7% русских - условно «кельтская» гаплогруппа R1b, которая доминирует у многих народов Западной Европы. Это наследие пребывания праславян в Центральной Европе. У 5% русских – «североевропейская» гаплогруппа I1 (I1a по старой классификации), распространенная в Скандинавии и Северной Европе. По-видимому, это потомки пресловутых «варягов». У 15% русских – «балканская» гаплогруппа I2 (ранее - I1b), характерная для народов Балканского полуострова. Это косвенное последствие славянской экспансии на Балканах в VI-IX вв., когда славянские пришельцы активно смешивались с местным автохтонным населением. Часть славян впоследствии ушла c Балкан на север и северо-восток (см. «Повесть временных лет»). У 4% - «восточно-средиземноморская» гаплогруппа J2, типичная для древних греков и еще более древних представителей минойской цивилизации. У русских это, скорее всего, составная часть «балканского наследия», а может - и более ранней эллинской колонизации Северного Причерноморья. У 6% русских представлена «афразийская» гаплогруппа E, частота которой у евреев, греков, балканских славян и южных итальянцев достигает 20% и более. Это снова «балканский след». В совокупности, потомки «византийских беженцев» (переселенцев с Балкан и Причерноморья) составляют среди русских более 25%. Мы видим, таким образом, (ка и ожидалось из ответа на второй вопрос) что гаплогруппы НИКАК не определяют "русскость". Чисто статистическое "преобладание" R1a1 равно 47 процентам. 53 процента русских НЕ ИМЕЮТ этой гаплогруппы и все равно являются русскими. Следовательно, если мы не ударимся в русофобию и не скажем, что 53 процента современных русских - "нечистые" (чего мы делать не станем), мы неизбежно вынуждены будем сделать еще раз тот же вывод, который был сделан выше - гаплогруппа не носит никаких специфически этноопределяющих черт, а этнос - неопределим через гаплогруппы. Вопрос 4: оправдана ли реконструкция этнических миграций и процессов на основе анализа современных гаплогрупп? Ответ: Нет. Проблем тут несколько. С большим скепсисом и скорее отрицательно можно воспринимать сообщения о "соотвествии распределения гаплогрупп историческим миграциям", а все "гаплогруппные" выводы о времени возникновения того или иного народа - с большой вероятностью принать по меньшей мере непроверяемыми и натянутыми. Почему? Потому что нужно не некритически цитировать "клесовых" ("ученый написал!") а ДУМАТЬ. Прежде всего - своей головой, используя логику и полученное образование. Во-первых, исторические миграции тех или иных народов в дописьменное время являются не аксиомой. а предметом научных споров и дискуссий - и в плане того, являлись ли носители той или иной археологической культуры представителями определенного этноязыковового единства (с точностью мы не никогда не сможем этого сказать - горшки и топоры не говорят), и в плане точности соответствия археологической культуры - определенному этносу или этнокультурному единству. Археологические культуры - это комплекс предметов материальной культуры, вещей, их остатков, вернее. С точки зрения археолога 30 века, на всей территории России, Европы. США в 21 веке будет существовать единая, с локальными вариациями, археологическая культура "шин, пластиковых пакетов, бутылок, банок и компьютеров", в которой различия наций по их материальной кльтуре видны не будут. Хотя мало оснований подразумевать подобную глобализацию в древности, стоит помнить о том. что чем древнее археологическая культура, тем размытей ее этническая принадлежность. Таким образом сразу ставится под сомнение тезис "скелет такой то, несомненно индоевропейский.. имеет такую то гаплогруппу". То что он "несомненно" индоевропейский, а не затесавшийся сюда инородец, принявший уклад этого племени, еще доказать надо. А при крайне нерепрезентативной выборке (согласитесь, десяток или сотня захоронений на огромную область - это не выборка, или, по крайней мере выборка, по репрезентативности несравнимая с современной выборкой среди живущих людей) это делается практически равновероятным, т.е неопределенным. Во вторых, проследить такие миграции по гаплогруппам крайне проблематично из-за того, что (в случае с индоевропейцами например) среди множества археологических культур бронзового века - "кухни пра-этносов", распространен погребальный обряд трупосожжения в различных его вариациях, то же самое на некоторых территориях и в веке железном. ДНК материал носителей этих культур безвозвратно утерян и мы ровным счетом ничего не можем сказать об их гаплогруппах. Таким образом, тут "гаплогрупповый" подход не работает - огромные территории и целые тысячелетия выпадают из его сферы напрочь, лишая нас возможности построить сколько-нибудь преемственную цепочку и так крайне скудного и нерепрезентативного по сути материала. В третьих, мутации в Y-хромосме происходят случайно и могут как увеличиваться так и "уменьшаться" (или возвращаться к первоначальному, исходному коду). Это указывает на то, что вы не можете определить время и место возникновения той или иной "группы" по причине и постоянных миграций, которые могли быть и по суше или морю, и мутаций, происходящих в течении всей жизни той или иной исторической человеческой общности. В четвертых, теоретически может быть подвергнута сомнению сама точность исследования современного распространения гаплогрупп - т.е. того исходного, экстраполируемого на прошлое, материала (трудность с определением этнической принадлежности лиц. проходящих тест - кто мешает вам назваться русским или белоруссом?). При относительно небольших выборках и спорных вопроса типа "русский-украинец" (фактически вольный вопрос самоназвания) это крайне важно и может решать все. Иначе говоря, перенесение современной дистрибуции наборов гаплогрупп в прошлое - неоправдан, а раз так, то и нельзя делать выводы об этничности тех или иных человеческих останков, выдавая их только лишь на этом основании за чьих-то предков. Это натянуто и поверхностно. Подводя итог всему описанному. - Ссылки на гаплогруппы как "безусловный показатель этничности" несостоятельны.- Гаплогруппы - это инструменты исследования, статистически значимые маркеры. Реальные же, - объективно наблюдаемые Y-хромосомы не несут значимой этнической информации в генокоде.- Русские (как и немцы, финны и тюрки и др) существуют и уникальны не за счет гаплогрупп.- Любой народ имеет не одну а две-три и более гаплогрупп в своем наборе, при этом нет оснований делать одних "менее чистыми" а других - "более чистыми", по причине описанной тезисом выше.- Мы априори не можем реконструировать по гаплогруппам достоверные миграции. а тем более - дату происхождения (возникновения_ того или иного современного этноса. Таким образом можете смело выбрасывать клесовские сочинения и сказки в ту же помойку, где у нас лежат Фоменко, Левашов и Чудинов. Автор: http://14-sonde.livejournal.com/ merjamaa.ru Что такое гаплогруппа? Анатолий Клёсов 20 родов человечества: какую метку носит в себе каждый? Почему о гаплогруппах нельзя читать в Википедии? Как работает ДНК-генеалогия? Сколько мутаций генов было на протяжении развития человеческого рода? Каким 200-250 тысяч лет назад был первопредок человека? Чем различаются 20 основных ДНК – 20 родов человечества? Имеют ли отношение к гаплогруппе этносы, нации, религии? Как получилось, что Пушкин – носитель гаплогруппы R1a? Откуда в Африке столько представителей гаплогруппы R1b? Могут ли на пути рода встречаться разные расы? Почему расы продолжают держаться раздельно? Существует ли русский геном? Анатолий Клёсов – основатель ДНК-генеалогии, доктор химических наук, вице-президент американской фармацевтической компании Galectin Therapeutics рассказывает, как меняется антропология, но остаётся неизменной гаплогруппа. Анатолий Клесов: Во-первых, когда Вы спрашиваете про гаплогруппы, не открывайте Википедию. Потеря времени, и получаете неверную информацию. Дело в том, что Википедия - это замечательный источник, но тогда, когда Вы ищете информацию установленную, скажем, сколько цифр в числе "пи", тут нет вопросов, или определение треугольника. Вот такие вещи давно отлажены и состыкованы к консенсусу, для таких вещей Википедия. Когда Вы хотите смотреть по гаплогруппам, сразу скажу, пишут там это люди, которые ничего в этом не понимают. Люди, которые думают, возможно, что понимают, но на самом деле ничего не понимают, руководствуются непонятно чем, там идет просто свалка информации абсолютно пустой и искаженной. Дело в том, что для этого придется для объяснения ответить на вопрос, что такое гаплогруппа и род, и почему это совершенно не связано, это такие перпендикулярные понятия. Скажем, этнос, национальность, партийность, профессия, все эти вещи, на самом деле, в той же мере не связаны с понятием гаплогруппы и рода. Дело в том, что такое гаплогруппа? И этому есть научное объяснение, есть научные критерии. Когда стали заниматься ДНК, это, я думаю, что на слуху это понятие, даже люди не знают детали, что это такое, но знают, что ДНК - это нечто в нас, в организме, которое определяет наши и антропологию, и внешний вид, и цвет волос, зачастую поведение, короче все характеристики, которые мы выражаем, к друг другу проявляем, это все заложено в ДНК. ДНК - гигантская молекула, там миллиарды звеньев в этой ДНК, и она каждое поколение переплетается, половина идет от мамы, половина от папы, и поэтому вносит вклад и с мужской, женской стороны, и опять же образуется новое переплетение. ДНК имеет вид такой двойной спирали. Одна часть спирали от мамы, другая от папы, вот она двойная спираль в какой-то степени с этим тоже связана, а потом следующие дети опять идут от новой мамы, папы переплетения, поэтому каждая ветка - изменение. Однако, часть ДНК зовется Y-хромосомой, по-научному говоря, мужская половая хромосома, она именно половая, потому что определяет пол, она есть только у мужчин, у женщин ее нет, поэтому сейчас археологи, которые копают, находят мизинчик, фрагмент мизинца, раньше, конечно, невозможно было определить мужской или женский. А тут извлекается ДНК, если это получается, и есть Y-хромосома, значит, это мужчина, мальчик, нет Y-хромосомы - женщина. Такие вещи теперь уже в археологии гораздо легче определять, то, что раньше практически не определялось. Так вот, дело в том, что у женщины Y-хромосомы нет, и она к полу не имеет никакого отношения, она вынашивает ребенка, мальчика или девочку. Вот собственно такая функция с точки зрения ДНК вещей. И вот ДНК получает человек, то есть мужчина, в данном случае, Y-хромосому получает он только от отца, у матери нет. А у отца от его отца, у отца от его отца и так миллионы лет без привлечений. От древних предков человека, древнейших предков человека идет одна и та же Y-хромосома. Более того, она по структуре почти идентична той, что есть у нас. Вот если взять шимпанзе из зоопарка, и взять у нее Y-хромосому, анализ на ДНК, то на 98% она такая же как у нас. А представляете себе, сколько времени прошло, то есть от древнего общего предка, когда-то был предок общий шимпанзе и человека, у него произошла развилка, и по одной ветке человек, а по другой шимпанзе, и продолжаются эти пути примерно 5 миллионов лет в одну сторону и 5 миллионов лет в другую сторону, то есть нашу шимпанзе отделяет 10 миллионов лет, обратно к предку и туда, напрямую мерить нельзя естественно. И за 10 миллионов лет 98% Y-хромосом осталось без изменений, поэтому здесь данные надежные, и их можно определять, ДНК изучается. И поскольку такое почти неизменное, хромосома передается, Y-хромосома, понятно, что изменений от отца к сыну немного. Действительно, во всей Y-хромосоме всего лишь одна мутация происходит в поколение, то есть за 22 года, математически возьмем примерно поколение, всего лишь на всю Y-хромосому. И поэтому вот такая одна мутация за поколение, каких-то 1000 поколений, 25000 лет, всего лишь 1000 мутаций образованы, что элементарно определяется современными методами, можно каждый из них пересчитать, и каждый уже пересчитан и пронумерован. И оказалось, что если исходить от, примерно, 200000 лет назад, это был общий предок Хомо Сапиенс, человек разумный, данные могут быть такие - 200000-250000 лет назад жил первый предок человека, а до этого были уже предки более глубокие, более архаичные, у них была другая антропология, другая форма грудной клетки, они уже не считаются Хомо Сапиенс, человек разумный. Так оказалось, что есть 20 линий основных ДНК, за эти 200000 лет они разошлись, вот 20 линий, это называли 20 родов человеческих. И каждая линия определяется одной конкретной мутацией, которая есть только у этой линии. Тут слово "линия" можно неправильно понять, не линия, а куст на самом деле. В основе каждой линии есть точка, то есть человек, от него куча потомков, от другой линии, от него свой куст потомков, а у третьего свой. Таких 20 кустов - это все человечество, и в каждом своя метка, которой нет ни у какого другого куста, поэтому эта метка определяет к какому кусту любой из нас принадлежит. Вот это есть гаплогруппа, вот этот куст, он же род. Почему род? То же самое определение, в основе был предок, патриарх, от него куст образовался. Слово "род", оно в быту имеет несколько понятие расплывчатое, скажем так. Я, например, из рода Клёсовых, а до этого, фамилий не было когда, род же продолжал быть, поэтому можно сказать, я из рода моего дедушки, прадедушки, а я из рода князя Голицына. Но до этого тоже не было князей, поэтому род, он смещается, какое понятие в род вкладывают, то и вкладывают, это в быту. Но это ничего не противоречит тому, что есть в науке, потому что в науке это все один большой род, но распадается на много мелких подродов, вплоть до семей, бытовых ячеек человечества. Поэтому, повторяю, гаплогруппа - это один куст, и возраст гаплогруппы часто, причем, бывает, что куст, он потом расходится на разные кусты, некоторые из которых более массивные, и они охватывают большую часть населения, бывает сотни миллионов людей в одном кусте. И вот куст, бывает, выделяют один, который позже образовался, но очень массивный, а другие почти все вымерли из предыдущего куста, поэтому гаплогруппой называют этот куст, который очень массивный. Вот сейчас принято условное определение, допустим, что гаплогруппа - это куст, в котором не менее 100 миллионов человек, это род, в основе этого рода тоже был один человек. Причем, понимаете, что к национальности не имеет никакого отношения, потому что гаплогруппа образовалась по разным данным, некоторые молодые 20 тысяч лет назад, некоторые 40 тысяч лет назад, 60 тысяч лет назад, а самые старые 200 тысяч лет назад. Конечно, понятие "национальность" никак не корреспондируется с понятием "гаплогруппа", национальности совсем недавно образовались по историческим меркам. Этносы совсем недавно тоже. Этнос - это общность опять же территории, языка и места проживания, ну да, территория и есть место проживания, языка, и зачастую культурные вещи, зачастую и религия вкладывает в понятие "этнос". Скажем, этнос есть, общая большая группа славян, а этносы разные. Этнос поляков, скажем, не есть этнос русских. Этнос украинцев, этнос белорусов может вполне, потому что язык немножко различается, опять же у них различие языка и территории, и культурных признаков. Поэтому этносы, национальности, религия, партийность, к гаплогруппе, к роду не имеют никакого отношения, но бывает, что так получилось, что эта группа образовалась, она живет в одном месте, и образовался вот этот этнос как раз в этих рамках, и тогда это, может, вполне корреспондируется с гаплогруппой, нельзя исключить такого. Например, есть баски, у них 90% одна и та же гаплогруппа, R1B. И есть этнос басков, то есть в этом случае этнос и гаплогруппа, они почти симпатны, почти накладываются друг на друга. Дело в том, что раса, она очень гибкое понятие, давайте вспомним Пушкина, такой заезжий пример. У Пушкина гаплогруппа R1A, это мы узнали по его детям, по его внукам, правнукам, с этим уже вопросом ясно. Более того, Пушкин происходит из такого не самого главного, но дворянского рода, у него предок давний был воевода Радша, которого называют иногда Рача, и он служил у Александра Невского. У него как раз R1A, классическая русская титульная, если угодно, гаплогруппа по численности. И он R1A, но мы знаем, что он не гройный, не в малой степени, он, говоря научным языком, метис, то есть у него антропология, раса, у него уже там где-то на стыке между европеоидной расой и негроидной, африканской. А, допустим, он женился бы на опять же негритянке, и у него дети были бы еще более в черную сторону, а те бы женились еще, то есть 2-3 поколения, и было бы уже не отличить от чернокожих африканцев, а гаплогруппа была бы R1A, Пушкины. Вот пример, что R1A, а это негроидные. В Африке живут все гаплогруппы, не знаю, R1A пока еще не встречалась, но R1B миллионы человек, то есть те, которые в свое время пришли не в Европу, а выбрали свободу, Южную Африку, своего миграционного пути, они расселились в районе Камеруна и Чады, и там миллионы человек, они говорят на языках местных банту, но они по гаплогруппе R1B. То же самое, раса черная, а R1B. Поэтому раса меняется, повторяю, буквально в 2-3 поколения. У меня есть коллекция фотографий, скажем, одна из них, хакас. Хакасы, они монголоидные, сибирская народность, и он, хакас, чисто монголоидный по лицу, женился на блондинке москвичке, так случилось, и пошли дети, у детей внуки. И у меня фотография внучки, они уже москвички, как я понимаю, и они играют где-то в песочнице, совершенно белокурые блондинки, от русских не отличить, а дедушка или прадедушка был монголоид полный. Поэтому здесь антропология меняется в момент. Вы уедете, повторяю, хотя всем уже ясно, уедете к американским индейцам, 2-3 поколения и не отличить, а гаплогруппа останется. Поэтому обычное понятие, спрашивают, как так может быть, R1A на Алтае, там монголоиды живут, такого не может быть, то есть люди не понимают, что это разные вещи. Род, который идет с древних времен, на этом пути рода могут быть какие угодно вариации в расе, нет никакой здесь корреляции. По той же причине, что в свое время древние арии, они прошли по территории всей Русской равнины, прошли дальше по южным территориям, прошли через Среднюю Азию и оставили много потомков среди киргизов. Среди киргизов любой, кто там жил, был или проезжал, знает, что есть много голубоглазые, светловолосые киргизы, совсем другие, он киргиз, и поэтому здесь опять же антропология расы, разрез глаз и прочее никак не связаны с R1A. Связаны, в каком смысле, если пойти в русскую деревню, конечно, вы там вряд ли встретите много австралийских аборигенов, американских индейцев или чернокожих африканцев, там все будут более-менее на одно лицо, они все будут светловолосые, типаж известен. Почему? Да потому что женятся на своих. Дело в том, что почему расы продолжают держаться раздельно? Потому что женятся, как правило, на своих. С чего бы крестьянин из русской деревни помчался жениться на дочери американского индейца? Такое бывает, но крайне редко. Во-первых, это так, чисто географически по возможностям всегда на своих. Далее, свои эталоны красоты, у монголов свой эталон красоты, плоское круглое лицо, они как луна, а у русского в эталон красоты это не входит, у него свои представления о красавицах, поэтому выбирают опять же по своим эталонам красоты. И вот так получается, что более или менее держится один. Если взять геном, кстати, вопрос спрашивают, а вот как геном, есть ли русский геном? Нет, русского генома нет, но, тем не менее, потому что русский геном опять же три разных, как правило, в основном, три разных рода его сложили, и каждый сделал вклады, где больше, где меньше, но если взять по всей России пройти, сделать усредненный геном, а потом по Африке усредненный геном, по Австралии, допусти, по аборигеном усредненный геном, то, конечно, он будет разный, поэтому вопрос всегда сводится к тому, а что мы хотим увидеть, и на какой вопрос можете этим ответить, и какими методами мы работаем, что мы, собственно говоря, делаем. Поэтому ответ, еще раз повторяю, важный, гаплогруппа не есть ни этнос, ни расы, ни национальность, и как я говорю, ни партийность, подчеркнуть, что это никак не связано, и не профессия, у профессии могут быть разные рода. Это следует различать, потому что тысячи людей ошибаются, думая, что это связано с национальностями и расами. dostoyanieplaneti.ru Гаплокарты: ДНК-генеалогия с высоты птичьего полета Читатели Переформата, которых интересуют не только исторические и публицистические статьи, но и материалы по ДНК-генеалогии, наверное, обратили внимание, что в начале 2016 года сайт Академии ДНК-генеалогии пополнился разделом «Гаплокарты». Таким, пока еще редко употребляющимся сокращением, автор этих строк назвал карты, на которых приведена статистика по основным Y-хромосомным гаплогруппам Евразии. Идея нанести на карту такие данные появилась, очевидно, с того самого момента, как они начали поступать в более-менее значимых количествах, то есть с конца 1990-х – начала 2000-х годов. Сейчас в книгах, журналах и Сети можно найти огромное количество карт на любой вкус и цвет, что в той или иной мере касаются Y-ДНК. Казалось бы, что нового может дать очередная их версия? Ответу на этот вопрос посвящен краткий обзор карт и таблиц, которые размещены в указанном разделе.  Вначале следует объяснить, чем не устраивают существующие карты. Прежде всего, при всем разнообразии, среди них нет таких, что в полной мере могли бы дать наиболее полный обзор распределения Y-гаплогрупп по регионам и этническим группам, где привлекались бы последние данные с максимально доступной на сегодняшний день репрезентативностью и разрешением по филогении. Имеющиеся в наличии карты и таблицы такой связной картины не дают. Они либо ограничиваются какой-либо конкретной гаплогруппой и регионом, либо используют устаревшую номенклатуру, имеют недостаточное разрешение по филогении, а зачастую опираются на выборки с низкой статистической значимостью. Например, таблица, размещенная в Википедии, предлагает информацию об Y-гаплогруппах голландцев по выборке из 27 (sic!) гаплотипов, со ссылками на публикации 2000 и 2004 гг. По миллиардным Индии и Китаю нет даже этого. Альтернативой подобной попгенетической архаике можно было бы считать карты и таблицы с проектов FTDNA, но их составители зачастую бросаются в другую крайность – настолько сильно дробят участников на категории и подкатегории, что для восстановления общей картины по тому или иному этносу требуется приложить немало усилий. Наконец, нельзя не обойти вниманием, что часто приводимые в справочной литературе карты гаплогрупп, имитирующие карты плотности населения, грешат неоправданной детализацией, не соответствующей реальному уровню знания. Это почти точная аналогия попгенетическим датировкам, что рассчитываются с погрешностями в десятки процентов, но с точностью до четвертого знака после запятой. Бывают и откровенные курьезы, как, например, шлейф ветви R1b-V88, проходящий через одно из самых малонаселенных мест на Земле – нагорья Тибести и Энеди в Сахаре (см. ниже). Тот, кто знает, как работают компьютерные программы, рисующие такие карты, понимает, что это издержки алгоритма, но многие принимают его причуды за чистую монету и устраивают бурное обсуждение по поводу того или иного максимума на карте плотности. Вначале следует объяснить, чем не устраивают существующие карты. Прежде всего, при всем разнообразии, среди них нет таких, что в полной мере могли бы дать наиболее полный обзор распределения Y-гаплогрупп по регионам и этническим группам, где привлекались бы последние данные с максимально доступной на сегодняшний день репрезентативностью и разрешением по филогении. Имеющиеся в наличии карты и таблицы такой связной картины не дают. Они либо ограничиваются какой-либо конкретной гаплогруппой и регионом, либо используют устаревшую номенклатуру, имеют недостаточное разрешение по филогении, а зачастую опираются на выборки с низкой статистической значимостью. Например, таблица, размещенная в Википедии, предлагает информацию об Y-гаплогруппах голландцев по выборке из 27 (sic!) гаплотипов, со ссылками на публикации 2000 и 2004 гг. По миллиардным Индии и Китаю нет даже этого. Альтернативой подобной попгенетической архаике можно было бы считать карты и таблицы с проектов FTDNA, но их составители зачастую бросаются в другую крайность – настолько сильно дробят участников на категории и подкатегории, что для восстановления общей картины по тому или иному этносу требуется приложить немало усилий. Наконец, нельзя не обойти вниманием, что часто приводимые в справочной литературе карты гаплогрупп, имитирующие карты плотности населения, грешат неоправданной детализацией, не соответствующей реальному уровню знания. Это почти точная аналогия попгенетическим датировкам, что рассчитываются с погрешностями в десятки процентов, но с точностью до четвертого знака после запятой. Бывают и откровенные курьезы, как, например, шлейф ветви R1b-V88, проходящий через одно из самых малонаселенных мест на Земле – нагорья Тибести и Энеди в Сахаре (см. ниже). Тот, кто знает, как работают компьютерные программы, рисующие такие карты, понимает, что это издержки алгоритма, но многие принимают его причуды за чистую монету и устраивают бурное обсуждение по поводу того или иного максимума на карте плотности.  Итак, чтобы пройти между Сциллой скудости исходных данных и Харибдой сомнительной детализации, требуется корректная постановка задачи и ясное понимание того, насколько полную информацию могут предоставить существующие данные по Y-хромосомным гаплогруппам. Чтобы добиться поставленной цели, то есть дать общий и, по возможности, объективный обзор с максимально возможным охватом по географии и филогении, следует, во-первых, отказаться от чрезмерной детализации и наносить на карты только статистику по ключевым гаплогруппам с их привязкой по странам или регионам больших стран. Тем самым можно получить сопоставимые данные по всем доступным регионам, без перекоса в пользу более изученных. Излишняя перегруженность карты деталями, помимо того, делает ее трудной для восприятия. От идеи представить карты в виде контуров плотности, при всей их наглядности, также следует отойти, чтобы не возникало соблазна интерпретировать «фантомы», созданные компьютерными программами. Самой надежной и наглядной формой подачи на данный момент можно считать хорошо проверенные круговые диаграммы, размещенные в соответствующих местах. Следующим этапом, после формы представления, следует считать отбор исходных статистических данных. Во-первых, они должны быть репрезентативны, то есть отображать реальную статистику по тому или иному региону или этнической группе, без серьезных перекосов. Во-вторых, они должны содержать информацию по всем гаплогруппам, задействованным в обзоре, либо должны позволять извлечь такую информацию из гаплотипов, не типировавшихся на снипы. И, в-третьих, желательно иметь как можно более свежие данные, чтобы избежать ошибок типирования, что были нередки на начальном этапе сбора информации. Как оказалось, значительная часть материала, собранного и опубликованного в самых цитируемых работах попгенетиков, не проходит по какому-либо из этих критериев, а то и по всем сразу. Этих недостатков лишены данные с географических ДНК-проектов, опубликованные на сайте компании FTDNA, но их охват по регионам очень неравномерен. Например, по Азии репрезентативные данные имеются только для армян, грузин и арабов из стран Персидского залива. Недостающие данные можно получить из судебно-медицинских выборок, опубликованных в специализированных журналах, а также в базе данных YHRD. Хотя в них, как правило, не определяют снипы, отнесение можно с достаточно высокой степенью надежности оценить по гаплотипам в 17- и 23-маркерных форматах, принятых экспертами-криминалистами в качестве стандарта. Методика работы с этими наборами хорошо отработана, и ошибки типирования в последние 3-4 года стали большой редкостью. В ряде случаев, этим данным отдавалось предпочтение даже по некоторым странам, хорошо представленным в проектах FTDNA, если требовалась привязка к конкретным регионам. Например, сводная статистика по русским была взята из полевой выборки 2008 года, чтобы иметь представление о географии гаплогрупп в пределах России. Ссылки на оригинальные работы и данные в табличной форме по каждой из диаграмм приведены в таблицах, размещенных рядом с картами. Пришлось пока отказаться от картирования Индии, точнее ее самой населенной северной части, потому что имеющиеся данные, хотя и исчисляются сотнями гаплотипов, не дают желаемой связной картины. В публикациях в обилии представлены различные этнические и социальные группы индийцев, но без соотнесения с демографией. Насколько они отображают ситуацию в целом по региону, судить сложно. К примеру, можно ли считать репрезентативной выборку русских, если она состоит из 50 человек с фамилией «Иванов», 50 – с фамилией «Клёсов» и 50 – с фамилией «Рожанский»? Может быть, да, а может быть, нет. Без отдельной проверки вывод сделать нельзя, что мы и наблюдаем с индийцами. Выборки, собранные без дробления на касты и племена, пока имеются только по тамилам с юга Индии и мусульманам из Бангладеш и Пакистана. По другим странам Азии таких проблем не возникало. Насколько можно судить, информация по гаплогруппам Азии в таком объеме публикуется впервые. Карты для Африки, Океании и коренного населения Америки пока не готовились из-за недостатка данных, а также из-за сравнительно небольшого набора гаплогрупп, присутствующих у живущих там народов. На этом первую часть обзора, а именно постановку задачи и описание исходных данных, можно считать завершенной. Во второй части предлагаю краткую характеристику генеалогических линий, представленных на гаплокартах. Гаплогруппа A1 Встречается крайне редко, а потому включена в раздел «прочие. Тем не менее, в Западной Европе живут носители загадочной гаплогруппы А1а, которой была посвящена заметка на Переформате. Гаплогруппа A1b найдена у арабов Персидского залива и Ливана, где представлена ветвью A1b1b-М32, пик распространения которой приходится на Эфиопию и нилотские народы Восточной Африки. Помимо арабов, единичные гаплотипы той же ветви отмечены у армян, в Казахстане, на Русской равнине и Британских островах. Очевидно, за пределы Восточной Африки субклад М32 распространился как минорная ветвь в миграциях народов – носителей галогруппы E1b1b1-М35, со стороны Африканского Рога, в том числе и путем работорговли. Гаплогруппа В Также из категории «прочие». Встречается за пределами Африки реже, чем гаплогруппа А. Обнаружена у арабов и хазарейцев из Афганистана. Вероятно, маркирует потомков рабов, вывезенных арабскими купцами из Восточной Африки. Следует отметить, что рабство в странах Ближнего Востока отличалось от рабства в Новом Свете XVI-XIX веков тем, что в нем не было столь выраженной сегрегации между рабами и хозяевами. В течение нескольких поколений потомки раба, если у него была семья, становились равноправными членами сообщества, поскольку ислам в его самой строгой форме отрицает сословное деление для его последователей. При удачном стечении обстоятельств человек, некогда купленный на невольничьем рынке, мог достичь самого высокого социального статуса, как показывают примеры мамлюкских султанов в Египте и Палестине. Гаплогруппа С Одна из древнейших и самых разветвленных гаплогрупп. Присутствует повсеместно, кроме Африки. Найдена у европейцев эпохи палеолита (Костёнки), мезолита (Испания) и раннего неолита (Венгрия), но в настоящее время ее в Европе настолько мало, что в выборке европейцев она входит в категорию «прочие». В Центральной и Восточной Азии, напротив, это одна из основных гаплогрупп, что представлена почти исключительно субкладом С2-М217, который расходится на большое количество ветвей с общим предком, жившим около 34000 лет назад. Родственный ему, и более древний «костенковский» субклад С1-F3393 рассеян от Европы до Океании. В выборках, представленных на гаплокартах, его ветви в значимых количествах присутствуют у японцев и филиппинцев на уровне 3-5%, а также найдены в древней ДНК из Красноярского края и с северо-востока Китая. Гаплогруппа DE Является предковой к гаплогруппам D и E, характеризуется цепочкой в 40 снипов в формате YFull. Единственный подтвержденный представитель – араб из Сирии. Сообщения из ранних статей о находках DE в Африке и на Тибете пока сложно подтвердить или опровергнуть из-за высокой вероятности «недотипирования», что часто имело место на начальном этапе сбора информации по Y-ДНК. Возможно, среди нескольких тысяч гаплотипов из полевых выборок Китая и Юго-Восточной Азии есть вероятность найти представителей чрезвычайно редких линий, восходящих непосредственно к DE, но из-за отсутствия данных по снипам их пока невозможно распознать. Гаплогруппа D Практически не встречается за пределами Восточной и Юго-Восточной Азии, где является одной из принципиальных генеалогических линий у тибетцев и японцев. Расходится на несколько далеко отстоящих ветвей с общим предком, жившим около 46000 лет назад, из которых субклад D-M64.1 ограничен почти исключительно Японским архипелагом и восходит, очевидно, к его древним обитателям из культуры Дзёмон (шнуровой керамики, в переводе с японского). Большой перевес гаплогруппы D на Тибете создается за счет одной из ветвей субклада D-P47, не имеющей пока своего снипа, но легко узнаваемой по редкому значению DYS392=7. Ее предок жил сравнительно недавно – около 4800 лет назад. Та же ветвь присутствует и у других народов Восточной Азии, но намного реже. Остальные ветви гаплогруппы D встречаются очень редко, и в отсутствие данных по снипам ее носителей распознать весьма сложно. Гаплогруппа E Наряду с J, O и R, входит в четверку «супер-гаплогрупп», число носителей которых исчисляется сотнями миллионов, а география захватывает целые континенты. Для гаплогруппы Е этот континент – Африка, хотя из рассмотрения гаплогрупп DE и D следует, что ее корни находятся вне Африки. На Черном Континенте представлены все основные субклады этой древней (около 54000 лет до предка) гаплогруппы, из которых только один, E1b1b1-M35, вышел за его пределами в значимых количествах. Соответственно, на гаплокартах отмечена статистика только по М35, а остальные внесены в категорию «прочие». Субклад E-M35 в Евразии расходится на 2 основные ветви, Е-М78 и Е-Z287, что имеют «возраст» около 10000 и 12000 лет, соответственно. В свою очередь, они дробятся на большое количество дочерних ветвей, из которых можно выделить E-V13, специфическую для Европы и начавшую свой рост около 3700 лет назад, как многие другие генеалогические линии Европы, прошедшей через демографический кризис на рубеже III и II тысячелетий до н.э. С другой стороны, находки ископаемых гаплотипов Е-V13 и E-M78 с датировками около 7000 лет назад в Испании и Венгрии говорят о том, что представители этих ветвей стали заселять Европу еще со времен раннего неолита. Гаплогруппа F Подтвержденных носителей этой гаплогруппы, не относящихся к дочерним GHIJK, в настоящее время известно крайне мало. Это единичные гаплотипы из Южной и Юго-Восточной Азии, к которым, возможно, добавится какое-то количество гаплотипов из выборок по этим регионам, не типированных пока на снипы. Для программы-предиктора они неотличимы от тех, что принадлежат к гаплогруппам G или H. Сопоставимое количество носителей гаплогруппы F было найдено в ископаемой ДНК в Европе, в том числе в останках 39000-летней давности из Румынии. Такой результат вполне ожидаем, если принять во внимание, что в гаплогруппы, дочерние к F, входит подавляющее большинство современных жителей Евразии, Океании и Америки. Гаплогруппа G Современные представители этой гаплогруппы имеют общего предка, жившего около 25000 лет назад, что заметно меньше, чем у рассмотренных выше линий. На гаплокарте дана статистика по самому населенному ее субкладу G2a-P15, что имеет «возраст» около 17000 лет и рассеян от Западной Европы до Центральной Азии. Несколько лет назад большим сюрпризом стали находки этой гаплогруппы в ископаемой ДНК Европы, где сейчас доля G2a невелика. В настоящее время, когда число образцов ископаемой Y-ДНК перевалило за 100, можно считать вполне установленным фактом то, что это была одна из основных линий Европы эпохи неолита. О распределении G2a в Азии той же эпохи пока нет информации, но, судя по современным данным с Кавказа и из Ирана, ее представители жили и там. Статистика по другим, меньшим по численности и более локальным по своей географии ветвям гаплогруппы G, собрана в категории «прочие». Это субклады G1 и G2b, рассеянные с низкой частотой в Передней и Средней Азии. Обе эти редкие ветви найдены также у евреев-ашкенази, что предполагает присутствие гаплогрупп G1 и G2b на Ближнем Востоке до начала диаспоры евреев. Гаплогруппа H Несмотря на раннее время расхождения на ветви (около 48000 лет до предка), эта гаплогруппа имеет довольно компактный ареал, ограниченный в основном Южной Азией. В Европе встречается, как правило, у цыган и их потомков, принесшим ее из Индии. Однако в ископаемой ДНК из Европы времен неолита были обнаружены представители ныне крайне редкой гаплогруппы Н2-Р96, что предполагает более широкую географию Н в прошлом. Гаплогруппа I Находится в родстве с распространенной на Ближнем Востоке и в Средиземноморье гаплогруппой J, с которой образует сводную гаплогруппу IJ, характеризуемую цепочкой из 64 снипов в формате YFull, но связана с Европой, где, очевидно, зародилась и начала расходиться около 27000 лет назад. В Азии встречается очень редко, а потому отнесена на гаплокарте Азии в категорию «прочие». На гаплокарте Европы, в силу особой значимости в этногенезе народов Европы, дана статистика по трем принципиальным дочерним гаплогруппам: I1-M253, I2a1-P37.2 и I2a2-M436. На сегодняшний день пока не найдено ни одного надежно подтвержденного гаплотипа из родительской гаплогруппы I, в том числе и в ископаемой ДНК. Перечисленные выше дочерние гаплогруппы охватывают абсолютное большинство современных носителей гаплогруппы I, за исключением очень редких I2b-L415 и I2c-L596, что помещены в категорию «прочие». Одна из ветвей I2c, а именно Y16419, компактно располагается в Восточном Причерноморье, где ее доля доходит до 5% в выборке по Восточной Грузии. Сравнительно небольшое, около 3500 лет назад, время жизни ее предка и находки родственных гаплотипов в Европе эпохи мезолита и раннего неолита дают основание считать, что эта линия имеет европейское происхождение. Гаплогруппа I1 Долгое время для этой гаплогруппы, рост которой начался около 4300 лет назад, постулировали североевропейское или даже скандинавское происхождение. Однако из гаплокарты Европы следует, что ее география намного шире, и охватывает весь континент, за исключением юга Балканского и Апеннинского полуостровов. Самые рано отошедшие ветви I1 распространены в Центральной и Восточной Европе, но почти не захватывают Скандинавский полуостров, что мало согласуется со скандинавской гипотезой. Находка I1 в ископаемой ДНК из Венгрии времен неолита (культура линейно-ленточной керамики) также можно считать доводом в пользу континентального происхождения этой гаплогруппы. Однако пока нет возможности установить, в каком месте Европы и в составе каких народов начался ее рост после прохождения бутылочного горлышка на рубеже III и II тысячелетий до н.э. Гаплогруппа I2a1 История этой гаплогруппы уходит ко временам палеолита (около 18500 лет до предка), но в настоящее время подавляющее большинство ее носителей принадлежит к ветви CTS10228, рост которой начался в эпоху античности, около 2300 лет назад. Высокий численный перевес и значительный процент этой ветви среди народов юго-востока Европы создали иллюзию балканского происхождения всей гаплогруппы I2, что, судя по ее филогении и данным древней ДНК, вряд ли соответствует действительности. Все остальные ветви гаплогруппы I2a1, кроме уже упомянутой CTS10228, рассеяны по Западной Европе. Одна из них, PF4188, является основной генеалогической линией жителей Сардинии, но, как и в случае с восточноевропейской ветвью CTS10228, ее предок жил сравнительно недавно – на рубеже нашей эры. В настоящее время пока сложно сказать с определенностью, где зародилась гаплогруппа I2a1 и каковы были ее пути при заселении Европы. Гаплогруппа I2a2 Время жизни предка I2a2 совпадает с тем, что оценено для I2a1, но, в отличие от последней, в ней нет таких многолюдных линий, как CTS10228. Она расходится на несколько примерно равных по численности ветвей, доля которых достигает 6-7% в выборках из Дании, Нидерландов и Англии. Наряду с I2a1 и I2c, носителей этой гаплогруппы идентифицировали в древней ДНК из различных частей Европы. Несмотря на небольшую численность, гаплогруппа I2a2 имеет широкий ареал, и в качестве минорной линии присутствует у всех народов Русской равнины, доходя до Афганистана и уйгуров из оазисов Синьцзяна. Возможно, какая-то часть ее носителей приняла участие в миграциях ариев, но по численности они сильно уступали представителям гаплогруппы R1a, и большинство их линий пресеклось. Гаплогруппа J Следующая из гаплогрупп-«тяжеловесов», как правило, сейчас редко рассматривается как единое целое. В базах данных и публикациях отдельно идет статистика по дочерним гаплогруппам J1 и J2. В современных выборках пока не найдено гаплотипов, которые можно считать подтвержденными представителями родительской ветви J. По данным древней ДНК, возможно, к ней принадлежал человек, живший около 7200 лет назад на территории современной Карелии, но из-за плохой сохранности ДНК многие ее фрагменты не прочитаны, что оставляет возможность отнесения либо к J1, либо к J2. Гаплогруппа J1 Является одной из основных генеалогических линий на Ближнем Востоке и на Кавказе, где представлена преимущественно субкладами YSC0000234 (около 4600 лет до предка) и Z1842 (около 6000 лет до предка), соответственно. Другие ветви встречаются заметно реже, но довольно сильно разбросаны по географии. К одной из них, F6304, вероятно, принадлежал человек, живший на территории Грузии около 13200 лет назад. Судя по этим данным, а также исходя из времени жизни предка всех современных представителей J1 (около 18500 лет назад), эта гаплогруппа с самых ранних времен занимала территории к югу от Большого Кавказского хребта, но более точно локализовать ее прародину пока не удается. В популярных изданиях гаплогруппу J1 нередко называют основной у евреев, но это не совсем верно. В репрезентативной выборке евреев-ашкенази она находится на третьем месте, немного уступая гаплогруппам E и J2. Гаплогруппа J2 Одна из самых старых (около 28000 лет до предка) и самых разветвленных генеалогических линий Евразии. Проходит широкой полосой от Пиренейского полуострова на западе до верховьев Хуанхэ на востоке, составляя значительную долю у народов Средиземноморья, Кавказа, Передней и Южной Азии. Многие из дочерних ветвей J2 имеют выраженную привязку к тем или иным регионам и этносам, что позволяет возводить их к миграциям разных эпох, от неолита до раннего Средневековья. Находки ископаемой ДНК из гаплогруппы J2 пока не очень многочисленны, но по ним можно судить о ее широкой географии уже в древние времена. Это Грузия 9600 лет назад, Турция 8400 лет назад, Венгрия 7000 и 3200 лет назад, Армения 3100 лет назад и Алтай начала нашей эры. Гаплогруппа K Подобно гаплогруппе F, в Евразии найдено очень мало подтвержденных представителей К, кто не входит в дочерние сводные гаплогруппы LT и NOP. Однако значительное количество таких линий существует в Австралии и Океании. В их числе минорные гаплогруппы M и S, с выделением которых в самостоятельные таксономические единицы явно поторопились. Можно ожидать, что в представленных на гаплокарте выборках из примыкающей к Океании Юго-Восточной Азии обнаружатся новые представители гаплогруппы К, но в отсутствие данных по снипам их распознать невозможно, и они, скорее всего, оказались на настоящий момент в списках гаплогрупп N, O или Q. В данных по ископаемой ДНК имеется пока всего один представитель предковой гаплогруппы К, но это самый древний на сегодняшний день человек, у которого была расшифрована Y-хромосома. Он жил в Западной Сибири около 47000 лет назад, а его останки (бедренную кость) нашли близ впадения Ишима в Иртыш, откуда и знакомое многим имя – человек из Усть-Ишима. По сумме косвенных данных, родиной гаплогруппы К можно считать Центральную, Восточную или Юго-Восточную Азию, но локализовать ее более точно пока сложно. Гаплогруппа L Входит в сводную гаплогруппу LT, в составе которой имеет 32 общих снипа с гаплогруппой T в формате YFull. Время жизни предка современных представителей гаплогруппы L оценивается как примерно 23000 лет назад. В выборках из Южной Азии ее доля достигает 7-15%, в остальных регионах встречается редко, потому на гаплокарте Европы помещена в категорию «прочие». Локальный максимум гаплогруппы L отмечен среди нескольких народов Кавказа. Он достигается за счет молодых ветвей Y6247 (около 1000 лет до предка) и PH8 (около 2400 лет до предка). Гаплогруппа N Находится в близком родстве с гаплогруппой О, основной генеалогической линией Восточной Азии. Их общий предок из сводной гаплогруппы NO жил около 36500 лет назад. Так же, как и у жившего 15000 годами спустя предка современных представителей гаплогруппы N, их родину можно достаточно надежно поместить в Восточную Азию. В выборках из Китая и Монголии гаплогруппа N занимает третье место по численности, после О и С, а в древней ДНК времен неолита и бронзы с севера Китая она оказывается основной. Среди народов Дальнего Востока и Восточной Сибири, не представленных пока на гаплокарте, гаплогруппа N также одна из основных. В Западной Сибири и в Европе из всех ее ветвей присутствует почти исключительно довольно молодая (около 4500 лет до предка) ветвь L1026, появление которой связывают с распространением финно-угорских языков. В отличие от Китая, в Европе пока мало находок ископаемой ДНК из гаплогруппы N. Это два образца с Валдайской возвышенности (4500 и 1100 лет назад) и один из Венгрии (2900 лет назад). Гаплогруппа О Видимо, занимает первое место в мире по численности из всех гаплогрупп, поскольку доминирует в густонаселенных странах Восточной и Юго-Восточной Азии. Расходится на большое количество ветвей, филогения и география которых находятся в процессе уточнения. По этой причине, а также чтобы не перегружать карту, на гаплокарте Азии приведена статистика по всей гаплогруппе, без разделения на субклады. Помимо Восточной Азии, гаплогруппа О составляет значительную долю у народов, говорящих на языках австронезийской семьи и расселившихся от Мадагаскара до Гавайских островов. На севере Китая найдено большое количество древней ДНК из различных ветвей гаплогруппы О, но, в отличие от современного населения, по числу находок она уступает гаплогруппам N и Q. Это дает дополнительный аргумент в пользу того, что первоначальный ареал гаплогруппы О охватывал юг современного Китая, а ее современное географическое распределение – результат миграций, начавшихся в эпоху неолита. Гаплогруппа Q Поскольку пока нет подтвержденных сведений о гаплотипах из родительской гаплогруппы Р, обзор продолжает гаплогруппа, которую можно считать безоговорочным лидером по распространению до эпохи Великих географических открытий. Ее ареал к началу Нового Времени занимал все северное полушарие, за исключением Африки, а также Южную Америку. Самое северное (Гренландия) и самое южное (Огненная Земля) места на Земле, где имеется постоянное население, были освоены представителями гаплогруппы Q. Однако, за исключением коренных жителей Америки и нескольких народов Сибири, гаплогруппа Q составляет незначительную долю у жителей Евразии. По абсолютной численности в Старом Свете, очевидно, лидирует ветвь М120, составляющая 2-4% в выборках ханьцев и найденная в древней ДНК на севере Китая. Больше всего различных линий этой очень разветвленной гаплогруппы отмечено в полосе от Алтая до Афганистана, в том числе и в ископаемой ДНК. Видимо, где-то в этом регионе около 32000 лет назад и жил ее предок. Локальные максимумы за пределами Центральной Азии отмечены у евреев-ашкенази (ветвь Y2200, около 1400 лет до предка), чеченцев (YP4000, 1200 лет) и жителей Скандинавии (L527 и L804, 2300-2100 лет), что косвенно говорит о некогда большой мобильности народов – носителей гаплогруппы Q. Гаплогруппа R Последняя по очереди и самая молодая из рассмотренных «супер-гаплогрупп». Ее предок жил около 31000 лет назад, очевидно, в Азии, как можно заключить из ее родства с азиатскими гаплогруппами NO и Q, а также из находки древней ДНК с палеолитической стоянки Мальта в Прибайкалье, датируемой 24000 годами назад. Следовательно, распространение гаплогруппы R, к началу Нового времени занимавшей Европу, Анатолию и Южную Азию, – результат древних миграций с востока на запад, детали которых пока неизвестны. Современные представители гаплогруппы R принадлежат к одной из трех дочерних гаплогрупп, R1a-M420, R1b-M343 и R2-M479, статистика по которым приведена на гаплокартах отдельно. Гаплогруппа R1a Предок этой многомиллионной гаплогруппы жил во времена палеолита, около 18000 лет назад, но абсолютное большинство современных представителей R1a ведет свой род от двух человек, живших намного позднее – около 5500 лет назад, когда начался рост субкладов Z282 и Z93. В свою очередь, они разошлись на многочисленные дочерние ветви, каждая со своей характерной географией и этническим составом. В общих чертах с ними можно ознакомиться по гаплокарте R1a, где указано распределение основных субкладов там, где позволял имеющийся материал. Гаплогруппа R1a найдена в многочисленных образцах древней ДНК, как правило, из археологических культур, связываемых с распространением индоевропейских языков – от культуры боевых топоров на западе до андроновской на востоке. След миграций древних ариев просматривается вплоть до Великой Китайской равнины и Таиланда, а на территории Индии их потомками является до 1/3 населения, преимущественно из ветвей L657 и Z2123. Очень редкие, рано отошедшие ветви YP4141, YP1272, YP1051 и L664 найдены пока в Европе и на Ближнем Востоке, в том числе ископаемый гаплотип из Карелии с датировкой около 7200 лет назад. Гаплогруппа R1b Датировки как предка всей гаплогруппы, так и доминирующих субкладов L51 и Z2103 практически совпадают с теми, что найдены в родственной гаплогруппе R1a. Однако ареал гаплогруппы R1b оказывается более разорванным, как можно заключить из составленной для нее гаплокарты. Западноевропейский субклад L51 и «анатолийский» Z2103 перекрываются в выборках из Центральной Европы и Южной Италии, но это, скорее, результат встречного движения с разных сторон, чем свидетельство центрально-европейского происхождения родительской для них ветви М269. Находки древней ДНК во многом соответствуют современному распределению этих субкладов (L51 – запад, Z2103 – восток), но, в отличие от заволжских степей (ямная и полтавкинская археологические культуры), носители гаплогруппы R1b появляются в Западной и Центральной Европе только в середине III тысячелетия до н.э. в захоронениях, относимых к культуре колоколовидных кубков. В пользу сравнительно позднего появления R1b в Европе косвенно свидетельствует и география рано отошедших ветвей V88 и M70, рассеянных по Центральной Азии, Ближнему Востоку и Центральной Африке (V88, см. рисунок выше). В отличие от родственной гаплогруппы R1a, пока не найдено каких-либо древних европейских ветвей R1b, причем на коммерческих выборках в десятки тысяч образцов в высоком разрешении. Гаплогруппа R2 Находится в тени более успешной в демографическом плане гаплогруппы R1, в том числе и потому, что ее ареал ограничен в основном Центральной и Южной Азией. Однако даже в Индии и Афганистане ее доля сравнительно невелика, и не превышает 10%. Восходит к предку, жившему около 11000 лет назад, видимо, в Центральной Азии, где живут представители самых рано отошедших ветвей R2. Вопреки распространенному мнению, гаплогруппа R2 у европейских цыган встречается очень редко, лишь в нескольких этнических группах. Родословные немногочисленных европейских представителей этой гаплогруппы восходят не столько к цыганам, сколько к евреям-ашкенази или потомкам уроженцев Ближнего Востока, разными путями попадавших в Европу. Гаплогруппа Т Входит в сводную гаплогруппу LT, но, в отличие от L, не имеет выраженной привязки к каким-либо регионам или этническим группам. В качестве минорной линии присутствует у всех народов Средиземноморья, Ближнего Востока и Передней Азии. Ввиду своей редкости, на гаплокарте Азии включена в категорию «прочие». Согласно полевым исследованиям, не включенным в гаплокарту, является одной из основных у некоторых народов Эфиопии. Найдена также в ископаемой ДНК из Германии времен культуры линейно-ленточной керамики (7500-6800 лет назад), что косвенно говорит о раннем ее распространении в ходе миграций эпохи раннего неолита. Какие-либо детали истории гаплогруппы Т пока не удается реконструировать, и она остается одной из самых загадочных гаплогрупп современных людей. В заключение, хотелось бы еще раз подчеркнуть, что представленный здесь обзор гаплогрупп не ставит своей целью дать исчерпывающую информацию по каждой из них. Его задача – очертить общую картину и отметить наиболее характерные черты. Дальнейшую информацию можно почерпнуть, изучая карты и специальную литературу, в том числе и в публикациях Академии ДНК-генеалогии. Приятного просмотра! Игорь Рожанский,кандидат химических наук Перейти к авторской колонке Итак, чтобы пройти между Сциллой скудости исходных данных и Харибдой сомнительной детализации, требуется корректная постановка задачи и ясное понимание того, насколько полную информацию могут предоставить существующие данные по Y-хромосомным гаплогруппам. Чтобы добиться поставленной цели, то есть дать общий и, по возможности, объективный обзор с максимально возможным охватом по географии и филогении, следует, во-первых, отказаться от чрезмерной детализации и наносить на карты только статистику по ключевым гаплогруппам с их привязкой по странам или регионам больших стран. Тем самым можно получить сопоставимые данные по всем доступным регионам, без перекоса в пользу более изученных. Излишняя перегруженность карты деталями, помимо того, делает ее трудной для восприятия. От идеи представить карты в виде контуров плотности, при всей их наглядности, также следует отойти, чтобы не возникало соблазна интерпретировать «фантомы», созданные компьютерными программами. Самой надежной и наглядной формой подачи на данный момент можно считать хорошо проверенные круговые диаграммы, размещенные в соответствующих местах. Следующим этапом, после формы представления, следует считать отбор исходных статистических данных. Во-первых, они должны быть репрезентативны, то есть отображать реальную статистику по тому или иному региону или этнической группе, без серьезных перекосов. Во-вторых, они должны содержать информацию по всем гаплогруппам, задействованным в обзоре, либо должны позволять извлечь такую информацию из гаплотипов, не типировавшихся на снипы. И, в-третьих, желательно иметь как можно более свежие данные, чтобы избежать ошибок типирования, что были нередки на начальном этапе сбора информации. Как оказалось, значительная часть материала, собранного и опубликованного в самых цитируемых работах попгенетиков, не проходит по какому-либо из этих критериев, а то и по всем сразу. Этих недостатков лишены данные с географических ДНК-проектов, опубликованные на сайте компании FTDNA, но их охват по регионам очень неравномерен. Например, по Азии репрезентативные данные имеются только для армян, грузин и арабов из стран Персидского залива. Недостающие данные можно получить из судебно-медицинских выборок, опубликованных в специализированных журналах, а также в базе данных YHRD. Хотя в них, как правило, не определяют снипы, отнесение можно с достаточно высокой степенью надежности оценить по гаплотипам в 17- и 23-маркерных форматах, принятых экспертами-криминалистами в качестве стандарта. Методика работы с этими наборами хорошо отработана, и ошибки типирования в последние 3-4 года стали большой редкостью. В ряде случаев, этим данным отдавалось предпочтение даже по некоторым странам, хорошо представленным в проектах FTDNA, если требовалась привязка к конкретным регионам. Например, сводная статистика по русским была взята из полевой выборки 2008 года, чтобы иметь представление о географии гаплогрупп в пределах России. Ссылки на оригинальные работы и данные в табличной форме по каждой из диаграмм приведены в таблицах, размещенных рядом с картами. Пришлось пока отказаться от картирования Индии, точнее ее самой населенной северной части, потому что имеющиеся данные, хотя и исчисляются сотнями гаплотипов, не дают желаемой связной картины. В публикациях в обилии представлены различные этнические и социальные группы индийцев, но без соотнесения с демографией. Насколько они отображают ситуацию в целом по региону, судить сложно. К примеру, можно ли считать репрезентативной выборку русских, если она состоит из 50 человек с фамилией «Иванов», 50 – с фамилией «Клёсов» и 50 – с фамилией «Рожанский»? Может быть, да, а может быть, нет. Без отдельной проверки вывод сделать нельзя, что мы и наблюдаем с индийцами. Выборки, собранные без дробления на касты и племена, пока имеются только по тамилам с юга Индии и мусульманам из Бангладеш и Пакистана. По другим странам Азии таких проблем не возникало. Насколько можно судить, информация по гаплогруппам Азии в таком объеме публикуется впервые. Карты для Африки, Океании и коренного населения Америки пока не готовились из-за недостатка данных, а также из-за сравнительно небольшого набора гаплогрупп, присутствующих у живущих там народов. На этом первую часть обзора, а именно постановку задачи и описание исходных данных, можно считать завершенной. Во второй части предлагаю краткую характеристику генеалогических линий, представленных на гаплокартах. Гаплогруппа A1 Встречается крайне редко, а потому включена в раздел «прочие. Тем не менее, в Западной Европе живут носители загадочной гаплогруппы А1а, которой была посвящена заметка на Переформате. Гаплогруппа A1b найдена у арабов Персидского залива и Ливана, где представлена ветвью A1b1b-М32, пик распространения которой приходится на Эфиопию и нилотские народы Восточной Африки. Помимо арабов, единичные гаплотипы той же ветви отмечены у армян, в Казахстане, на Русской равнине и Британских островах. Очевидно, за пределы Восточной Африки субклад М32 распространился как минорная ветвь в миграциях народов – носителей галогруппы E1b1b1-М35, со стороны Африканского Рога, в том числе и путем работорговли. Гаплогруппа В Также из категории «прочие». Встречается за пределами Африки реже, чем гаплогруппа А. Обнаружена у арабов и хазарейцев из Афганистана. Вероятно, маркирует потомков рабов, вывезенных арабскими купцами из Восточной Африки. Следует отметить, что рабство в странах Ближнего Востока отличалось от рабства в Новом Свете XVI-XIX веков тем, что в нем не было столь выраженной сегрегации между рабами и хозяевами. В течение нескольких поколений потомки раба, если у него была семья, становились равноправными членами сообщества, поскольку ислам в его самой строгой форме отрицает сословное деление для его последователей. При удачном стечении обстоятельств человек, некогда купленный на невольничьем рынке, мог достичь самого высокого социального статуса, как показывают примеры мамлюкских султанов в Египте и Палестине. Гаплогруппа С Одна из древнейших и самых разветвленных гаплогрупп. Присутствует повсеместно, кроме Африки. Найдена у европейцев эпохи палеолита (Костёнки), мезолита (Испания) и раннего неолита (Венгрия), но в настоящее время ее в Европе настолько мало, что в выборке европейцев она входит в категорию «прочие». В Центральной и Восточной Азии, напротив, это одна из основных гаплогрупп, что представлена почти исключительно субкладом С2-М217, который расходится на большое количество ветвей с общим предком, жившим около 34000 лет назад. Родственный ему, и более древний «костенковский» субклад С1-F3393 рассеян от Европы до Океании. В выборках, представленных на гаплокартах, его ветви в значимых количествах присутствуют у японцев и филиппинцев на уровне 3-5%, а также найдены в древней ДНК из Красноярского края и с северо-востока Китая. Гаплогруппа DE Является предковой к гаплогруппам D и E, характеризуется цепочкой в 40 снипов в формате YFull. Единственный подтвержденный представитель – араб из Сирии. Сообщения из ранних статей о находках DE в Африке и на Тибете пока сложно подтвердить или опровергнуть из-за высокой вероятности «недотипирования», что часто имело место на начальном этапе сбора информации по Y-ДНК. Возможно, среди нескольких тысяч гаплотипов из полевых выборок Китая и Юго-Восточной Азии есть вероятность найти представителей чрезвычайно редких линий, восходящих непосредственно к DE, но из-за отсутствия данных по снипам их пока невозможно распознать. Гаплогруппа D Практически не встречается за пределами Восточной и Юго-Восточной Азии, где является одной из принципиальных генеалогических линий у тибетцев и японцев. Расходится на несколько далеко отстоящих ветвей с общим предком, жившим около 46000 лет назад, из которых субклад D-M64.1 ограничен почти исключительно Японским архипелагом и восходит, очевидно, к его древним обитателям из культуры Дзёмон (шнуровой керамики, в переводе с японского). Большой перевес гаплогруппы D на Тибете создается за счет одной из ветвей субклада D-P47, не имеющей пока своего снипа, но легко узнаваемой по редкому значению DYS392=7. Ее предок жил сравнительно недавно – около 4800 лет назад. Та же ветвь присутствует и у других народов Восточной Азии, но намного реже. Остальные ветви гаплогруппы D встречаются очень редко, и в отсутствие данных по снипам ее носителей распознать весьма сложно. Гаплогруппа E Наряду с J, O и R, входит в четверку «супер-гаплогрупп», число носителей которых исчисляется сотнями миллионов, а география захватывает целые континенты. Для гаплогруппы Е этот континент – Африка, хотя из рассмотрения гаплогрупп DE и D следует, что ее корни находятся вне Африки. На Черном Континенте представлены все основные субклады этой древней (около 54000 лет до предка) гаплогруппы, из которых только один, E1b1b1-M35, вышел за его пределами в значимых количествах. Соответственно, на гаплокартах отмечена статистика только по М35, а остальные внесены в категорию «прочие». Субклад E-M35 в Евразии расходится на 2 основные ветви, Е-М78 и Е-Z287, что имеют «возраст» около 10000 и 12000 лет, соответственно. В свою очередь, они дробятся на большое количество дочерних ветвей, из которых можно выделить E-V13, специфическую для Европы и начавшую свой рост около 3700 лет назад, как многие другие генеалогические линии Европы, прошедшей через демографический кризис на рубеже III и II тысячелетий до н.э. С другой стороны, находки ископаемых гаплотипов Е-V13 и E-M78 с датировками около 7000 лет назад в Испании и Венгрии говорят о том, что представители этих ветвей стали заселять Европу еще со времен раннего неолита. Гаплогруппа F Подтвержденных носителей этой гаплогруппы, не относящихся к дочерним GHIJK, в настоящее время известно крайне мало. Это единичные гаплотипы из Южной и Юго-Восточной Азии, к которым, возможно, добавится какое-то количество гаплотипов из выборок по этим регионам, не типированных пока на снипы. Для программы-предиктора они неотличимы от тех, что принадлежат к гаплогруппам G или H. Сопоставимое количество носителей гаплогруппы F было найдено в ископаемой ДНК в Европе, в том числе в останках 39000-летней давности из Румынии. Такой результат вполне ожидаем, если принять во внимание, что в гаплогруппы, дочерние к F, входит подавляющее большинство современных жителей Евразии, Океании и Америки. Гаплогруппа G Современные представители этой гаплогруппы имеют общего предка, жившего около 25000 лет назад, что заметно меньше, чем у рассмотренных выше линий. На гаплокарте дана статистика по самому населенному ее субкладу G2a-P15, что имеет «возраст» около 17000 лет и рассеян от Западной Европы до Центральной Азии. Несколько лет назад большим сюрпризом стали находки этой гаплогруппы в ископаемой ДНК Европы, где сейчас доля G2a невелика. В настоящее время, когда число образцов ископаемой Y-ДНК перевалило за 100, можно считать вполне установленным фактом то, что это была одна из основных линий Европы эпохи неолита. О распределении G2a в Азии той же эпохи пока нет информации, но, судя по современным данным с Кавказа и из Ирана, ее представители жили и там. Статистика по другим, меньшим по численности и более локальным по своей географии ветвям гаплогруппы G, собрана в категории «прочие». Это субклады G1 и G2b, рассеянные с низкой частотой в Передней и Средней Азии. Обе эти редкие ветви найдены также у евреев-ашкенази, что предполагает присутствие гаплогрупп G1 и G2b на Ближнем Востоке до начала диаспоры евреев. Гаплогруппа H Несмотря на раннее время расхождения на ветви (около 48000 лет до предка), эта гаплогруппа имеет довольно компактный ареал, ограниченный в основном Южной Азией. В Европе встречается, как правило, у цыган и их потомков, принесшим ее из Индии. Однако в ископаемой ДНК из Европы времен неолита были обнаружены представители ныне крайне редкой гаплогруппы Н2-Р96, что предполагает более широкую географию Н в прошлом. Гаплогруппа I Находится в родстве с распространенной на Ближнем Востоке и в Средиземноморье гаплогруппой J, с которой образует сводную гаплогруппу IJ, характеризуемую цепочкой из 64 снипов в формате YFull, но связана с Европой, где, очевидно, зародилась и начала расходиться около 27000 лет назад. В Азии встречается очень редко, а потому отнесена на гаплокарте Азии в категорию «прочие». На гаплокарте Европы, в силу особой значимости в этногенезе народов Европы, дана статистика по трем принципиальным дочерним гаплогруппам: I1-M253, I2a1-P37.2 и I2a2-M436. На сегодняшний день пока не найдено ни одного надежно подтвержденного гаплотипа из родительской гаплогруппы I, в том числе и в ископаемой ДНК. Перечисленные выше дочерние гаплогруппы охватывают абсолютное большинство современных носителей гаплогруппы I, за исключением очень редких I2b-L415 и I2c-L596, что помещены в категорию «прочие». Одна из ветвей I2c, а именно Y16419, компактно располагается в Восточном Причерноморье, где ее доля доходит до 5% в выборке по Восточной Грузии. Сравнительно небольшое, около 3500 лет назад, время жизни ее предка и находки родственных гаплотипов в Европе эпохи мезолита и раннего неолита дают основание считать, что эта линия имеет европейское происхождение. Гаплогруппа I1 Долгое время для этой гаплогруппы, рост которой начался около 4300 лет назад, постулировали североевропейское или даже скандинавское происхождение. Однако из гаплокарты Европы следует, что ее география намного шире, и охватывает весь континент, за исключением юга Балканского и Апеннинского полуостровов. Самые рано отошедшие ветви I1 распространены в Центральной и Восточной Европе, но почти не захватывают Скандинавский полуостров, что мало согласуется со скандинавской гипотезой. Находка I1 в ископаемой ДНК из Венгрии времен неолита (культура линейно-ленточной керамики) также можно считать доводом в пользу континентального происхождения этой гаплогруппы. Однако пока нет возможности установить, в каком месте Европы и в составе каких народов начался ее рост после прохождения бутылочного горлышка на рубеже III и II тысячелетий до н.э. Гаплогруппа I2a1 История этой гаплогруппы уходит ко временам палеолита (около 18500 лет до предка), но в настоящее время подавляющее большинство ее носителей принадлежит к ветви CTS10228, рост которой начался в эпоху античности, около 2300 лет назад. Высокий численный перевес и значительный процент этой ветви среди народов юго-востока Европы создали иллюзию балканского происхождения всей гаплогруппы I2, что, судя по ее филогении и данным древней ДНК, вряд ли соответствует действительности. Все остальные ветви гаплогруппы I2a1, кроме уже упомянутой CTS10228, рассеяны по Западной Европе. Одна из них, PF4188, является основной генеалогической линией жителей Сардинии, но, как и в случае с восточноевропейской ветвью CTS10228, ее предок жил сравнительно недавно – на рубеже нашей эры. В настоящее время пока сложно сказать с определенностью, где зародилась гаплогруппа I2a1 и каковы были ее пути при заселении Европы. Гаплогруппа I2a2 Время жизни предка I2a2 совпадает с тем, что оценено для I2a1, но, в отличие от последней, в ней нет таких многолюдных линий, как CTS10228. Она расходится на несколько примерно равных по численности ветвей, доля которых достигает 6-7% в выборках из Дании, Нидерландов и Англии. Наряду с I2a1 и I2c, носителей этой гаплогруппы идентифицировали в древней ДНК из различных частей Европы. Несмотря на небольшую численность, гаплогруппа I2a2 имеет широкий ареал, и в качестве минорной линии присутствует у всех народов Русской равнины, доходя до Афганистана и уйгуров из оазисов Синьцзяна. Возможно, какая-то часть ее носителей приняла участие в миграциях ариев, но по численности они сильно уступали представителям гаплогруппы R1a, и большинство их линий пресеклось. Гаплогруппа J Следующая из гаплогрупп-«тяжеловесов», как правило, сейчас редко рассматривается как единое целое. В базах данных и публикациях отдельно идет статистика по дочерним гаплогруппам J1 и J2. В современных выборках пока не найдено гаплотипов, которые можно считать подтвержденными представителями родительской ветви J. По данным древней ДНК, возможно, к ней принадлежал человек, живший около 7200 лет назад на территории современной Карелии, но из-за плохой сохранности ДНК многие ее фрагменты не прочитаны, что оставляет возможность отнесения либо к J1, либо к J2. Гаплогруппа J1 Является одной из основных генеалогических линий на Ближнем Востоке и на Кавказе, где представлена преимущественно субкладами YSC0000234 (около 4600 лет до предка) и Z1842 (около 6000 лет до предка), соответственно. Другие ветви встречаются заметно реже, но довольно сильно разбросаны по географии. К одной из них, F6304, вероятно, принадлежал человек, живший на территории Грузии около 13200 лет назад. Судя по этим данным, а также исходя из времени жизни предка всех современных представителей J1 (около 18500 лет назад), эта гаплогруппа с самых ранних времен занимала территории к югу от Большого Кавказского хребта, но более точно локализовать ее прародину пока не удается. В популярных изданиях гаплогруппу J1 нередко называют основной у евреев, но это не совсем верно. В репрезентативной выборке евреев-ашкенази она находится на третьем месте, немного уступая гаплогруппам E и J2. Гаплогруппа J2 Одна из самых старых (около 28000 лет до предка) и самых разветвленных генеалогических линий Евразии. Проходит широкой полосой от Пиренейского полуострова на западе до верховьев Хуанхэ на востоке, составляя значительную долю у народов Средиземноморья, Кавказа, Передней и Южной Азии. Многие из дочерних ветвей J2 имеют выраженную привязку к тем или иным регионам и этносам, что позволяет возводить их к миграциям разных эпох, от неолита до раннего Средневековья. Находки ископаемой ДНК из гаплогруппы J2 пока не очень многочисленны, но по ним можно судить о ее широкой географии уже в древние времена. Это Грузия 9600 лет назад, Турция 8400 лет назад, Венгрия 7000 и 3200 лет назад, Армения 3100 лет назад и Алтай начала нашей эры. Гаплогруппа K Подобно гаплогруппе F, в Евразии найдено очень мало подтвержденных представителей К, кто не входит в дочерние сводные гаплогруппы LT и NOP. Однако значительное количество таких линий существует в Австралии и Океании. В их числе минорные гаплогруппы M и S, с выделением которых в самостоятельные таксономические единицы явно поторопились. Можно ожидать, что в представленных на гаплокарте выборках из примыкающей к Океании Юго-Восточной Азии обнаружатся новые представители гаплогруппы К, но в отсутствие данных по снипам их распознать невозможно, и они, скорее всего, оказались на настоящий момент в списках гаплогрупп N, O или Q. В данных по ископаемой ДНК имеется пока всего один представитель предковой гаплогруппы К, но это самый древний на сегодняшний день человек, у которого была расшифрована Y-хромосома. Он жил в Западной Сибири около 47000 лет назад, а его останки (бедренную кость) нашли близ впадения Ишима в Иртыш, откуда и знакомое многим имя – человек из Усть-Ишима. По сумме косвенных данных, родиной гаплогруппы К можно считать Центральную, Восточную или Юго-Восточную Азию, но локализовать ее более точно пока сложно. Гаплогруппа L Входит в сводную гаплогруппу LT, в составе которой имеет 32 общих снипа с гаплогруппой T в формате YFull. Время жизни предка современных представителей гаплогруппы L оценивается как примерно 23000 лет назад. В выборках из Южной Азии ее доля достигает 7-15%, в остальных регионах встречается редко, потому на гаплокарте Европы помещена в категорию «прочие». Локальный максимум гаплогруппы L отмечен среди нескольких народов Кавказа. Он достигается за счет молодых ветвей Y6247 (около 1000 лет до предка) и PH8 (около 2400 лет до предка). Гаплогруппа N Находится в близком родстве с гаплогруппой О, основной генеалогической линией Восточной Азии. Их общий предок из сводной гаплогруппы NO жил около 36500 лет назад. Так же, как и у жившего 15000 годами спустя предка современных представителей гаплогруппы N, их родину можно достаточно надежно поместить в Восточную Азию. В выборках из Китая и Монголии гаплогруппа N занимает третье место по численности, после О и С, а в древней ДНК времен неолита и бронзы с севера Китая она оказывается основной. Среди народов Дальнего Востока и Восточной Сибири, не представленных пока на гаплокарте, гаплогруппа N также одна из основных. В Западной Сибири и в Европе из всех ее ветвей присутствует почти исключительно довольно молодая (около 4500 лет до предка) ветвь L1026, появление которой связывают с распространением финно-угорских языков. В отличие от Китая, в Европе пока мало находок ископаемой ДНК из гаплогруппы N. Это два образца с Валдайской возвышенности (4500 и 1100 лет назад) и один из Венгрии (2900 лет назад). Гаплогруппа О Видимо, занимает первое место в мире по численности из всех гаплогрупп, поскольку доминирует в густонаселенных странах Восточной и Юго-Восточной Азии. Расходится на большое количество ветвей, филогения и география которых находятся в процессе уточнения. По этой причине, а также чтобы не перегружать карту, на гаплокарте Азии приведена статистика по всей гаплогруппе, без разделения на субклады. Помимо Восточной Азии, гаплогруппа О составляет значительную долю у народов, говорящих на языках австронезийской семьи и расселившихся от Мадагаскара до Гавайских островов. На севере Китая найдено большое количество древней ДНК из различных ветвей гаплогруппы О, но, в отличие от современного населения, по числу находок она уступает гаплогруппам N и Q. Это дает дополнительный аргумент в пользу того, что первоначальный ареал гаплогруппы О охватывал юг современного Китая, а ее современное географическое распределение – результат миграций, начавшихся в эпоху неолита. Гаплогруппа Q Поскольку пока нет подтвержденных сведений о гаплотипах из родительской гаплогруппы Р, обзор продолжает гаплогруппа, которую можно считать безоговорочным лидером по распространению до эпохи Великих географических открытий. Ее ареал к началу Нового Времени занимал все северное полушарие, за исключением Африки, а также Южную Америку. Самое северное (Гренландия) и самое южное (Огненная Земля) места на Земле, где имеется постоянное население, были освоены представителями гаплогруппы Q. Однако, за исключением коренных жителей Америки и нескольких народов Сибири, гаплогруппа Q составляет незначительную долю у жителей Евразии. По абсолютной численности в Старом Свете, очевидно, лидирует ветвь М120, составляющая 2-4% в выборках ханьцев и найденная в древней ДНК на севере Китая. Больше всего различных линий этой очень разветвленной гаплогруппы отмечено в полосе от Алтая до Афганистана, в том числе и в ископаемой ДНК. Видимо, где-то в этом регионе около 32000 лет назад и жил ее предок. Локальные максимумы за пределами Центральной Азии отмечены у евреев-ашкенази (ветвь Y2200, около 1400 лет до предка), чеченцев (YP4000, 1200 лет) и жителей Скандинавии (L527 и L804, 2300-2100 лет), что косвенно говорит о некогда большой мобильности народов – носителей гаплогруппы Q. Гаплогруппа R Последняя по очереди и самая молодая из рассмотренных «супер-гаплогрупп». Ее предок жил около 31000 лет назад, очевидно, в Азии, как можно заключить из ее родства с азиатскими гаплогруппами NO и Q, а также из находки древней ДНК с палеолитической стоянки Мальта в Прибайкалье, датируемой 24000 годами назад. Следовательно, распространение гаплогруппы R, к началу Нового времени занимавшей Европу, Анатолию и Южную Азию, – результат древних миграций с востока на запад, детали которых пока неизвестны. Современные представители гаплогруппы R принадлежат к одной из трех дочерних гаплогрупп, R1a-M420, R1b-M343 и R2-M479, статистика по которым приведена на гаплокартах отдельно. Гаплогруппа R1a Предок этой многомиллионной гаплогруппы жил во времена палеолита, около 18000 лет назад, но абсолютное большинство современных представителей R1a ведет свой род от двух человек, живших намного позднее – около 5500 лет назад, когда начался рост субкладов Z282 и Z93. В свою очередь, они разошлись на многочисленные дочерние ветви, каждая со своей характерной географией и этническим составом. В общих чертах с ними можно ознакомиться по гаплокарте R1a, где указано распределение основных субкладов там, где позволял имеющийся материал. Гаплогруппа R1a найдена в многочисленных образцах древней ДНК, как правило, из археологических культур, связываемых с распространением индоевропейских языков – от культуры боевых топоров на западе до андроновской на востоке. След миграций древних ариев просматривается вплоть до Великой Китайской равнины и Таиланда, а на территории Индии их потомками является до 1/3 населения, преимущественно из ветвей L657 и Z2123. Очень редкие, рано отошедшие ветви YP4141, YP1272, YP1051 и L664 найдены пока в Европе и на Ближнем Востоке, в том числе ископаемый гаплотип из Карелии с датировкой около 7200 лет назад. Гаплогруппа R1b Датировки как предка всей гаплогруппы, так и доминирующих субкладов L51 и Z2103 практически совпадают с теми, что найдены в родственной гаплогруппе R1a. Однако ареал гаплогруппы R1b оказывается более разорванным, как можно заключить из составленной для нее гаплокарты. Западноевропейский субклад L51 и «анатолийский» Z2103 перекрываются в выборках из Центральной Европы и Южной Италии, но это, скорее, результат встречного движения с разных сторон, чем свидетельство центрально-европейского происхождения родительской для них ветви М269. Находки древней ДНК во многом соответствуют современному распределению этих субкладов (L51 – запад, Z2103 – восток), но, в отличие от заволжских степей (ямная и полтавкинская археологические культуры), носители гаплогруппы R1b появляются в Западной и Центральной Европе только в середине III тысячелетия до н.э. в захоронениях, относимых к культуре колоколовидных кубков. В пользу сравнительно позднего появления R1b в Европе косвенно свидетельствует и география рано отошедших ветвей V88 и M70, рассеянных по Центральной Азии, Ближнему Востоку и Центральной Африке (V88, см. рисунок выше). В отличие от родственной гаплогруппы R1a, пока не найдено каких-либо древних европейских ветвей R1b, причем на коммерческих выборках в десятки тысяч образцов в высоком разрешении. Гаплогруппа R2 Находится в тени более успешной в демографическом плане гаплогруппы R1, в том числе и потому, что ее ареал ограничен в основном Центральной и Южной Азией. Однако даже в Индии и Афганистане ее доля сравнительно невелика, и не превышает 10%. Восходит к предку, жившему около 11000 лет назад, видимо, в Центральной Азии, где живут представители самых рано отошедших ветвей R2. Вопреки распространенному мнению, гаплогруппа R2 у европейских цыган встречается очень редко, лишь в нескольких этнических группах. Родословные немногочисленных европейских представителей этой гаплогруппы восходят не столько к цыганам, сколько к евреям-ашкенази или потомкам уроженцев Ближнего Востока, разными путями попадавших в Европу. Гаплогруппа Т Входит в сводную гаплогруппу LT, но, в отличие от L, не имеет выраженной привязки к каким-либо регионам или этническим группам. В качестве минорной линии присутствует у всех народов Средиземноморья, Ближнего Востока и Передней Азии. Ввиду своей редкости, на гаплокарте Азии включена в категорию «прочие». Согласно полевым исследованиям, не включенным в гаплокарту, является одной из основных у некоторых народов Эфиопии. Найдена также в ископаемой ДНК из Германии времен культуры линейно-ленточной керамики (7500-6800 лет назад), что косвенно говорит о раннем ее распространении в ходе миграций эпохи раннего неолита. Какие-либо детали истории гаплогруппы Т пока не удается реконструировать, и она остается одной из самых загадочных гаплогрупп современных людей. В заключение, хотелось бы еще раз подчеркнуть, что представленный здесь обзор гаплогрупп не ставит своей целью дать исчерпывающую информацию по каждой из них. Его задача – очертить общую картину и отметить наиболее характерные черты. Дальнейшую информацию можно почерпнуть, изучая карты и специальную литературу, в том числе и в публикациях Академии ДНК-генеалогии. Приятного просмотра! Игорь Рожанский,кандидат химических наук Перейти к авторской колонке Читайте другие статьи на Переформате: pereformat.ru Гаплогруппа I гаплогруппы I1 и I2 коренных европейцев народов Европы Если среди современных европейских народов Европы можно выделить несколько основных гаплогрупп (более 5%), это конечно же R1b, R1a, I1, N1, I2, G2 на севере и J, E1b1, T на юге Европы, то касательно населения древней Европы ситуация совсем иная.  Убежища последнего ледникового максимума Убежища последнего ледникового максимума
Происхождение гаплогруппы I Происхождение древних европейцев несомненно можно отождествлять с ДНК гаплогруппой I, отделившейся от пришедшей с Ближнего Востока Y гаплогруппой IJ и C6, которые были кроманьонцами.В популяционной генетике человека, Гаплогруппа I (M170, P19, M258) Y — хромосомная гаплогруппа, предположительно зародилась 20-25 тысяч лет назад на Балканском полуострове. Гаплогруппа I встречается почти у 1/5 европейского населения.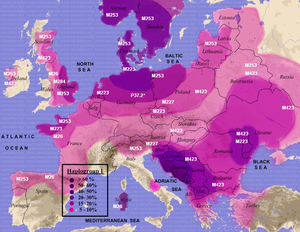 Некоторые исследователи полагают, что местом появления гаплогруппы I могла быть Граветтская культура, своего рода обитаемый "оазис", где люди переживали Последний ледниковый максимум, который произошел примерно 21 тысяч лет назад. Со времён прихода в Европу людей современного вида (кроманьонцев) около 40 тыс. лет назад и до индоевропейского вторжения около 5 тыс. лет назад, по всей видимости, гаплогруппа I составляла большинство населения Европы, возможно, существовало большее количество субклад, ныне исчезнувших, из которых ныне идентифицировано лишь две (I1 и I2). Строители западноевропейских мегалитов, вероятнее всего, принадлежали к гаплогруппе I2a. Некоторые исследователи полагают, что местом появления гаплогруппы I могла быть Граветтская культура, своего рода обитаемый "оазис", где люди переживали Последний ледниковый максимум, который произошел примерно 21 тысяч лет назад. Со времён прихода в Европу людей современного вида (кроманьонцев) около 40 тыс. лет назад и до индоевропейского вторжения около 5 тыс. лет назад, по всей видимости, гаплогруппа I составляла большинство населения Европы, возможно, существовало большее количество субклад, ныне исчезнувших, из которых ныне идентифицировано лишь две (I1 и I2). Строители западноевропейских мегалитов, вероятнее всего, принадлежали к гаплогруппе I2a. Этногеографическое распространение гаплогруппы I Наибольшее распространение гаплогруппы I имеется в 2 очагах распространения: первый это гаплогруппа I1 c центром в Скандинавских странах среди шведов, финнов, датчан и норвежцев.Второй это гаплогруппа I2 с центром на Балканах в Хорватии и Сербии. И гаплогруппа I1 и I2 распространены и в остальных странах Европы приблизительно в соотношении 15% в среднем. Гаплогруппа I1 Гаплогруппа I1 является коренной европейской гаплогруппой. Большинство современных носителей гаплогруппы I1 составляют носители германских языков индоевропейской семьи, хотя первоначально данная гаплогруппа была связана, по-видимому, не с индоевропейскими народами, а с догерманским субстратом (возможно Культура воронковидных кубков). I1 идентифицируется по меньшей мере 15 уникальными мутациями, что означает, что данная группа либо была совершенно изолирована в течение долгого периода (что маловероятно), либо пережила серьёзное «бутылочное горлышко» в сравнительно недавнем времени. Хотя первая мутация, отделившая I1 от I, могла произойти ещё 20 тысячелетий назад, все сегодняшние носители этой ДНК гаплогруппы происходят от одного мужчины, жившего не раньше, чем 5 тысячелетий назад. Это вполне совпадает со временем прихода на Скандинавию индоевропейцев, которые, как предполагается, уничтожали большую часть мужчин коренного населения или ставили их семьи в невыгодное демографическое положение. Так что выглядит вполне правдоподобным, что это вторжение пережил лишь один род коренных скандинавов (или, например, возможно, один мальчик), потомки которого в дальнейшем и составили гаплогруппу I1, которая таким образом стала достоверной меткой скандинавского п-ва, и складывавшихся на нем этнических образований в ту эпоху. Сегодня представителей этой группы находят почти всюду в Европе, некоторые субклады распространялись с германскими миграциями, единственным исключением может быть назван север европейской части России (Карелия, Вологда) и большая часть Поволжья, куда гаплогруппа I1 передалась через финнов. Гаплогруппа I2 Гаплогруппа I2 имеет 3 основных субклада с разным этногеографическим распространением в Европе:Субклад I2-M26 это одна из основных Y-хромосомных линий на Сардинии (достигает 40%) и у басков. С низкой частотой встречается также по всей юго-западной Европе: В Испании, Франции, Португалии, Италии.Субклад I2-M423 характерен для населения Балкан и Карпат, чаще всего наблюдается у динарийских славян (сербы, хорваты и боснийцы), а также в Молдове и на юго-западной Украине. Также с заметной частотой обнаруживается в Албании, северной Греции, Болгарии, Словакии, на восточной Украине, в Белоруссии и юго-западной России.Субклад I2-M223 характерен для северо-западной Европы и достигает максимума в Германии (Нижняя Саксония). Также встречается в Румынии, Молдавии и с низкой частотой в Восточной Европе. Гаплогруппы современных Европейцев после индоевропейской экспансии и Великого переселения народов  Среди современных этнических европейцев распространены следующие гаплогруппы в порядке убывания (включая южную и восточную Европу): гаплогруппа R1b1a2 (33.71%),гаплогруппа R1a1 (22.21%), гаплогруппа I1 (8.39%), гаплогруппа I2a (7.97%), гаплогруппа E1b1b1 (6.77%),гаплогруппа J2 (6.48%), гаплогруппа N1c1 (5.82%), гаплогруппа G2a3 (3.92%), гаплогруппа I2b (2.20%) и с низкой частотой встречаются гаплогруппы T (1.82%), Q1a (1,32%) и J1 (0,85%). Гаплогруппы с суммарной частотой менее 0.5% на диаграмме не приведены.Данные проценты гаплогрупп среди европейцев учитывают численность коренного населения европейских стран с населением свыше 1 млн. человек и среднюю частоту гаплогрупп по ним из нижеприведенной таблицы, суммарная погрешность не превышает 5%. Среди современных этнических европейцев распространены следующие гаплогруппы в порядке убывания (включая южную и восточную Европу): гаплогруппа R1b1a2 (33.71%),гаплогруппа R1a1 (22.21%), гаплогруппа I1 (8.39%), гаплогруппа I2a (7.97%), гаплогруппа E1b1b1 (6.77%),гаплогруппа J2 (6.48%), гаплогруппа N1c1 (5.82%), гаплогруппа G2a3 (3.92%), гаплогруппа I2b (2.20%) и с низкой частотой встречаются гаплогруппы T (1.82%), Q1a (1,32%) и J1 (0,85%). Гаплогруппы с суммарной частотой менее 0.5% на диаграмме не приведены.Данные проценты гаплогрупп среди европейцев учитывают численность коренного населения европейских стран с населением свыше 1 млн. человек и среднюю частоту гаплогрупп по ним из нижеприведенной таблицы, суммарная погрешность не превышает 5%.  Данные гаплогруппы с разной частотой от 0-95% встречаются среди немцев, испанцев, итальянцев, греков, поляков, венгров, французов, албанцев, англичан, ирландцев, шотландцев, португальцев, швейцарцев, чехов, словаков, сербов, хорватов, румынов, белорусов, украинцев, датчан, шведов, норвежцев, финнов, русских, эстонцев, латышей, литовцев и др. Генетика европейцев Сами гаплогруппы не несут генетической информации, т.к. генетическая информация находиться в аутосомах- первых 22 пар хромосом. Посмотреть распространение генетических компонент в Европе можно здесь. Гаплогруппы являются всего лишь метками давно минувших дней, на заре формирования современных народов. Карта распространения гаплогрупп в Европе из диссертации О. Балановского(только для представленных более 35%) Продолжение по Y-гаплогруппам европейцев I, R1a, R1b и N1c1Продолжение по Y-гаплогруппам европейцев G, J, E1b1b1 и TПродолжение по Y-гаплогруппам европейцев Q, L, C и др.Информация по мито-гаплогруппам европейцев Подробное филогенетическое дерево Y гаплогруппдДНК из ископаемых останков различных эпохСкачать диссертацию О. Балановского о распределении Y и мт-ДНК гаплогрупп [зеркало] |
haplogroup.narod.ru
|











